

Autocitrullination of human peptidyl arginine deiminase type 4 regulates protein citrullination during cell activation
- PMID: 20201080
- PMCID: PMC2951335
- DOI: 10.1002/art.27439
Autocitrullination of human peptidyl arginine deiminase type 4 regulates protein citrullination during cell activation
Abstract
Objective: To address mechanisms that control the activity of human peptidyl arginine deiminase type 4 (PAD-4).
Methods: PAD-4 autocitrullination was determined by anti-modified citrulline immunoblotting, using purified recombinant and endogenous PAD-4 from activated human primary neutrophils and cell lines expressing PAD-4. The citrullination sites in PAD-4 were determined by mass spectrometry. Mechanisms of autocitrullination-induced inactivation and the functional consequences of autocitrullination in PAD-4 polymorphic variants were addressed using purified components and cell lines expressing PAD-4 wild-type, PAD-4 mutant, and PAD-4 polymorphic variants relevant to rheumatoid arthritis (RA).
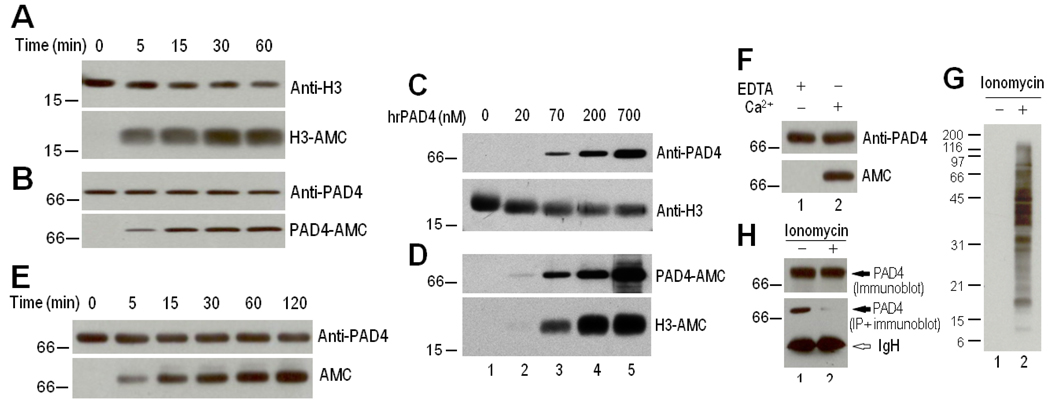
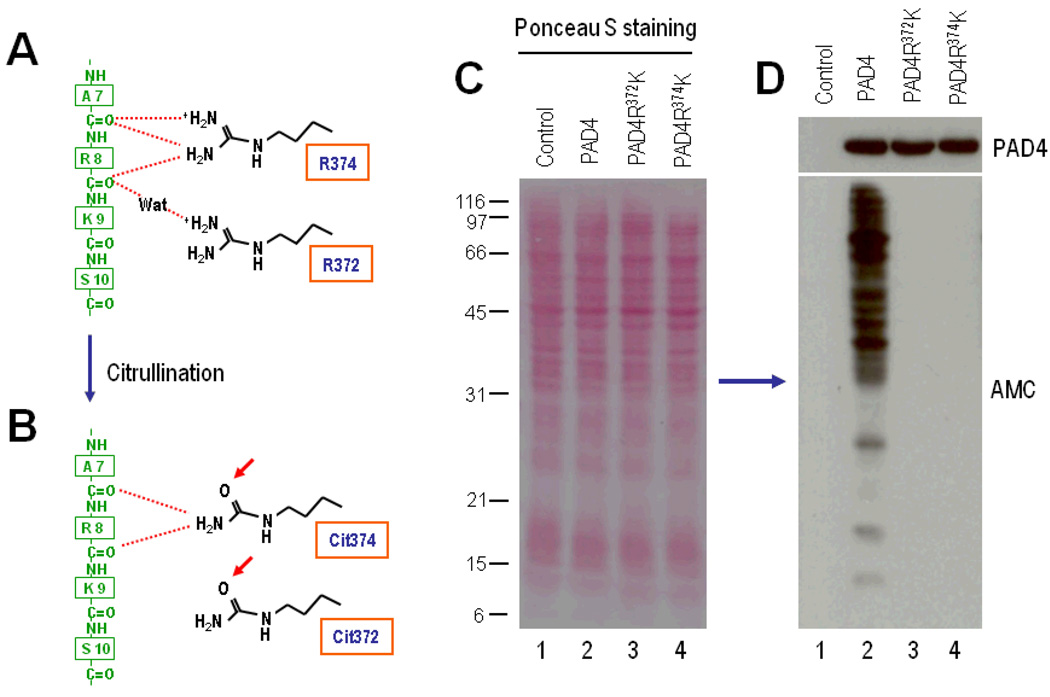
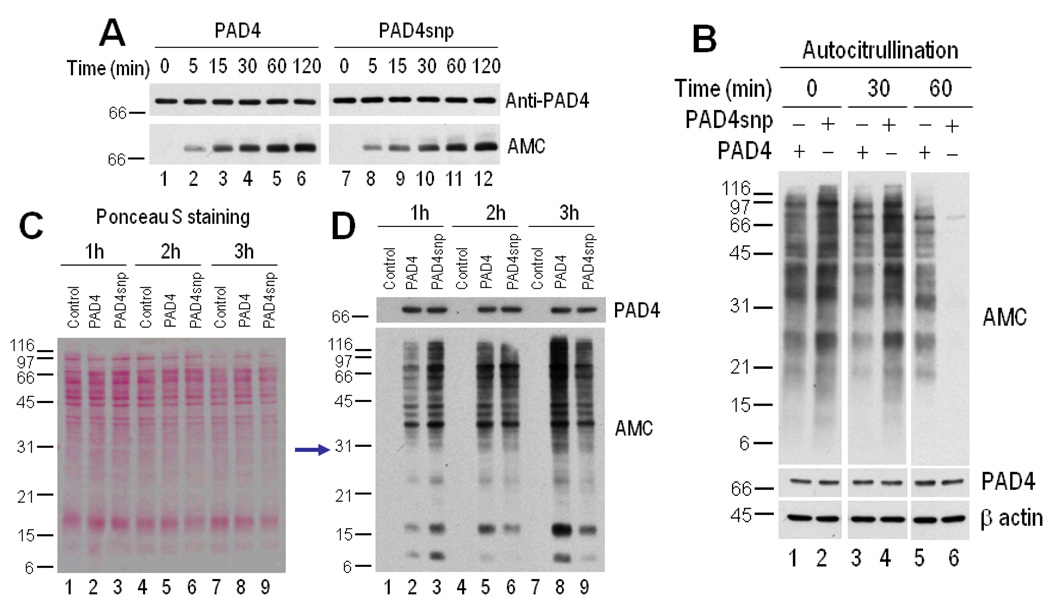
Results: PAD-4 is autocitrullinated in vitro and during activation of primary cells and cell lines expressing PAD-4. Interestingly, this modification inactivated the function of the enzyme. The efficiency of inactivation differed among genetically defined PAD-4 variants relevant to RA. PAD-4 was citrullinated at 10 sites, which are clustered into 3 distinct regions, including a cluster of arginines around the active site cleft where Arg-372 and -374 were identified as the potential autocitrullination targets that inactivate the enzyme. Autocitrullination also modified the structure of PAD-4, abrogating its recognition by multiple rabbit antibodies, but augmenting its recognition by human anti-PAD-4 autoantibodies.
Conclusion: Our findings suggest that autocitrullination regulates the production of citrullinated proteins during cell activation, and that this is affected by structural polymorphisms in PAD-4. Autocitrullination also influences PAD-4 structure and immune response.
Figures



Similar articles
-
Citrulline Not a Major Determinant in the Recognition of Peptidylarginine Deiminase 2 and 4 by Autoantibodies in Rheumatoid Arthritis.Arthritis Rheumatol. 2020 Sep;72(9):1476-1482. doi: 10.1002/art.41276. Epub 2020 Jul 14. Arthritis Rheumatol. 2020. PMID: 32255561 Free PMC article.
-
Peptidylarginine deiminase 2, 3 and 4 have distinct specificities against cellular substrates: novel insights into autoantigen selection in rheumatoid arthritis.Ann Rheum Dis. 2012 Jan;71(1):92-8. doi: 10.1136/ard.2011.151712. Epub 2011 Aug 21. Ann Rheum Dis. 2012. PMID: 21859690 Free PMC article.
-
Defining the role of Porphyromonas gingivalis peptidylarginine deiminase (PPAD) in rheumatoid arthritis through the study of PPAD biology.Ann Rheum Dis. 2015 Nov;74(11):2054-61. doi: 10.1136/annrheumdis-2014-205385. Epub 2014 May 26. Ann Rheum Dis. 2015. PMID: 24864075 Free PMC article.
-
PAD enzymes in rheumatoid arthritis: pathogenic effectors and autoimmune targets.Nat Rev Rheumatol. 2020 Jun;16(6):301-315. doi: 10.1038/s41584-020-0409-1. Epub 2020 Apr 27. Nat Rev Rheumatol. 2020. PMID: 32341463 Review.
-
Citrullination by peptidylarginine deiminase in rheumatoid arthritis.Ann N Y Acad Sci. 2007 Jun;1108:323-39. doi: 10.1196/annals.1422.034. Ann N Y Acad Sci. 2007. PMID: 17893996 Review.
Cited by
-
Activation of PAD4 in NET formation.Front Immunol. 2012 Nov 29;3:360. doi: 10.3389/fimmu.2012.00360. eCollection 2012. Front Immunol. 2012. PMID: 23264775 Free PMC article.
-
Citrullinome of Porphyromonas gingivalis Outer Membrane Vesicles: Confident Identification of Citrullinated Peptides.Mol Cell Proteomics. 2020 Jan;19(1):167-180. doi: 10.1074/mcp.RA119.001700. Epub 2019 Nov 21. Mol Cell Proteomics. 2020. PMID: 31754044 Free PMC article.
-
An unbiased proteomic analysis of PAD4 in human monocytes: novel substrates, binding partners and subcellular localizations.Philos Trans R Soc Lond B Biol Sci. 2023 Nov 20;378(1890):20220477. doi: 10.1098/rstb.2022.0477. Epub 2023 Oct 2. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37778379 Free PMC article.
-
PAD2-mediated citrullination of Fibulin-5 promotes elastogenesis.Matrix Biol. 2021 Aug;102:70-84. doi: 10.1016/j.matbio.2021.07.001. Epub 2021 Jul 15. Matrix Biol. 2021. PMID: 34274450 Free PMC article.
-
A cyclic peptide toolkit reveals mechanistic principles of peptidylarginine deiminase IV regulation.Nat Commun. 2024 Nov 11;15(1):9746. doi: 10.1038/s41467-024-53554-1. Nat Commun. 2024. PMID: 39528459 Free PMC article.
References
-
- Gelato KA, Fischle W. Role of histone modifications in defining chromatin structure and function. Biol Chem. 2008;389(4):353–363. - PubMed
-
- Tarcsa E, Marekov LN, Mei G, Melino G, Lee SC, Steinert PM. Protein unfolding by peptidylarginine deiminase. Substrate specificity and structural relationships of the natural substrates trichohyalin and filaggrin. J Biol Chem. 1996;271(48):30709–30716. - PubMed
-
- Doyle HA, Mamula MJ. Posttranslational modifications of self-antigens. Ann N Y Acad Sci. 2005;1050:1–9. - PubMed
-
- Rogers GE. Occurrence of citrulline in proteins. Nature. 1962;194:1149–1151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
