

Cocaine-induced neuroadaptations in glutamate transmission: potential therapeutic targets for craving and addiction
- PMID: 20201846
- PMCID: PMC5413205
- DOI: 10.1111/j.1749-6632.2009.05144.x
Cocaine-induced neuroadaptations in glutamate transmission: potential therapeutic targets for craving and addiction
Abstract
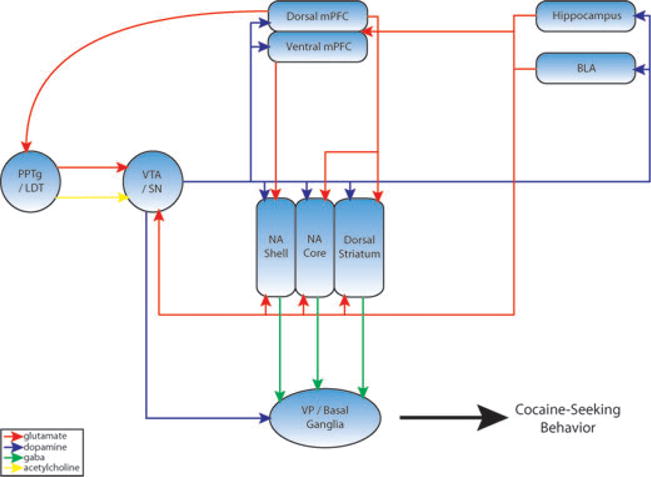
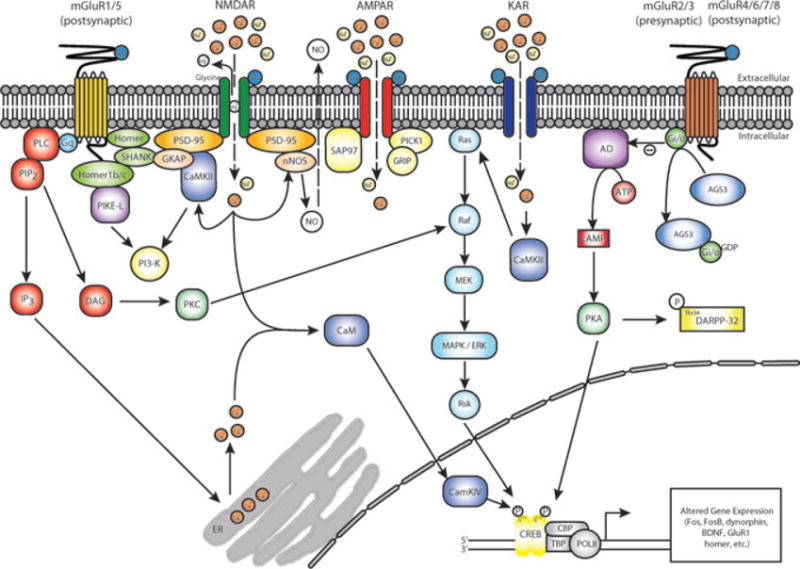
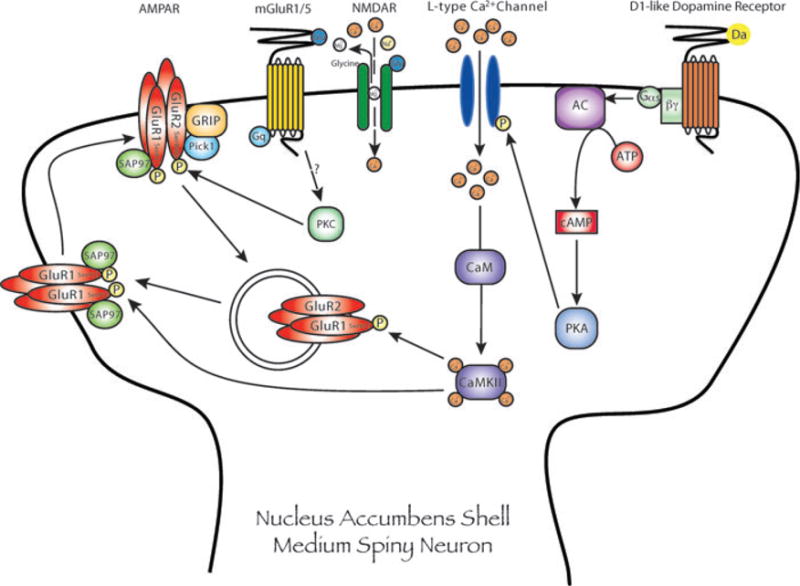
A growing body of evidence indicates that repeated exposure to cocaine leads to profound changes in glutamate transmission in limbic nuclei, particularly the nucleus accumbens. This review focuses on preclinical studies of cocaine-induced behavioral plasticity, including behavioral sensitization, self-administration, and the reinstatement of cocaine seeking. Behavioral, pharmacological, neurochemical, electrophysiological, biochemical, and molecular biological changes associated with cocaine-induced plasticity in glutamate systems are reviewed. The ultimate goal of these lines of research is to identify novel targets for the development of therapies for cocaine craving and addiction. Therefore, we also outline the progress and prospects of glutamate modulators for the treatment of cocaine addiction.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Glutamate: the new frontier in pharmacotherapy for cocaine addiction.CNS Neurol Disord Drug Targets. 2008 Nov;7(5):482-91. doi: 10.2174/187152708786927868. CNS Neurol Disord Drug Targets. 2008. PMID: 19128205 Review.
-
Long-term depression in the nucleus accumbens: a neural correlate of behavioral sensitization to cocaine.Nat Neurosci. 2001 Dec;4(12):1217-23. doi: 10.1038/nn757. Nat Neurosci. 2001. PMID: 11694884
-
Glutamate transmission in the nucleus accumbens mediates relapse in cocaine addiction.J Neurosci. 2000 Aug 1;20(15):RC89. doi: 10.1523/JNEUROSCI.20-15-j0006.2000. J Neurosci. 2000. PMID: 10899176 Free PMC article.
-
Cocaine sensitization and craving: differing roles for dopamine and glutamate in the nucleus accumbens.J Addict Dis. 2001;20(3):43-54. doi: 10.1300/J069v20n03_05. J Addict Dis. 2001. PMID: 11681592 Review.
-
Silent Synapse-Based Circuitry Remodeling in Drug Addiction.Int J Neuropsychopharmacol. 2016 Apr 29;19(5):pyv136. doi: 10.1093/ijnp/pyv136. Print 2016 May. Int J Neuropsychopharmacol. 2016. PMID: 26721952 Free PMC article. Review.
Cited by
-
Imbalances in prefrontal cortex CC-Homer1 versus CC-Homer2 expression promote cocaine preference.J Neurosci. 2013 May 8;33(19):8101-13. doi: 10.1523/JNEUROSCI.1727-12.2013. J Neurosci. 2013. PMID: 23658151 Free PMC article.
-
Effects of adenosine A2A receptor antagonists on cocaine-induced locomotion and cocaine seeking.Psychopharmacology (Berl). 2019 Feb;236(2):699-708. doi: 10.1007/s00213-018-5097-z. Epub 2018 Nov 3. Psychopharmacology (Berl). 2019. PMID: 30392131 Free PMC article.
-
Single Low Dose of Cocaine-Structural Brain Injury Without Metabolic and Behavioral Changes.Front Neurosci. 2021 Jan 22;14:589897. doi: 10.3389/fnins.2020.589897. eCollection 2020. Front Neurosci. 2021. PMID: 33584173 Free PMC article.
-
Drug-evoked synaptic plasticity in addiction: from molecular changes to circuit remodeling.Neuron. 2011 Feb 24;69(4):650-63. doi: 10.1016/j.neuron.2011.01.017. Neuron. 2011. PMID: 21338877 Free PMC article. Review.
-
Rac1 is essential in cocaine-induced structural plasticity of nucleus accumbens neurons.Nat Neurosci. 2012 Jun;15(6):891-6. doi: 10.1038/nn.3094. Nat Neurosci. 2012. PMID: 22522400 Free PMC article.
References
-
- Substance Abuse and Mental Health Services Administration. Results from the 2007 National Survey on Drug Use and Health. National Findings Office of Applied Studies; Rockville, MD: 2008.
-
- Hunt WA, Barnett LW, Branch LG. Relapse rates in addiction programs. J Clin Psychol. 1971;27:455–456. - PubMed
-
- O’Brien CP. A range of research-based pharmacotherapies for addiction. Science. 1997;278:66–70. - PubMed
-
- Carter BL, Tiffany ST. Cue-reactivity and the future of addiction research. Addiction. 1999;94:349–351. - PubMed
-
- de Wit H, Stewart J. Reinstatement of cocaine-reinforced responding in the rat. Psychopharmacology (Berl) 1981;75:134–143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
