

Quantal release of acetylcholine in mice with reduced levels of the vesicular acetylcholine transporter
- PMID: 20202084
- PMCID: PMC2866190
- DOI: 10.1111/j.1471-4159.2010.06657.x
Quantal release of acetylcholine in mice with reduced levels of the vesicular acetylcholine transporter
Abstract
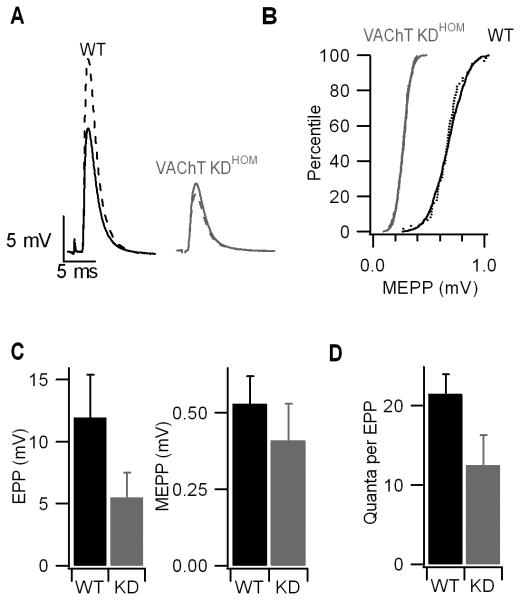
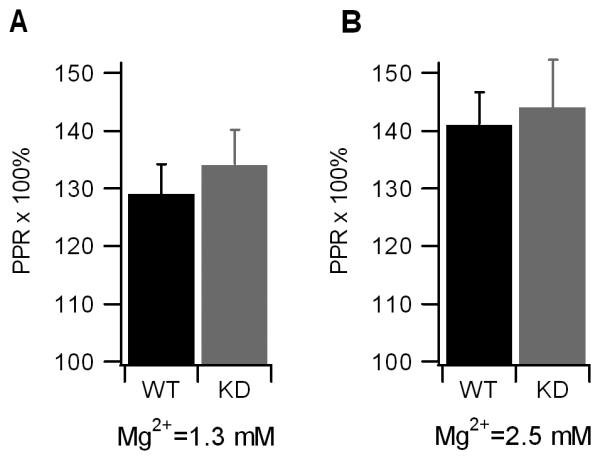
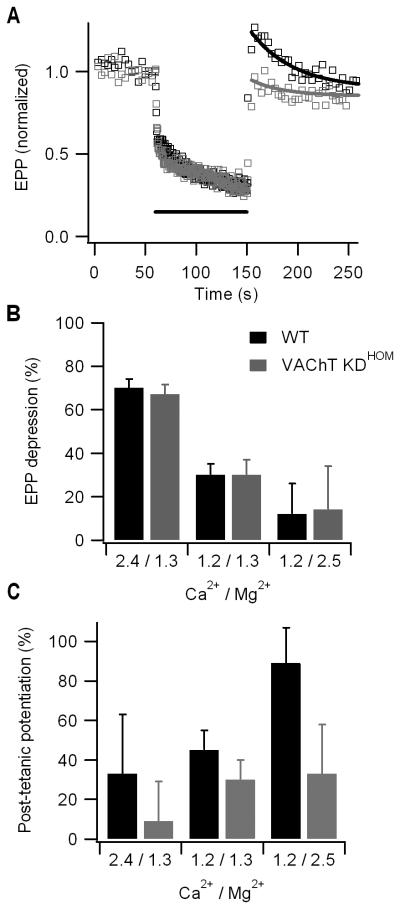
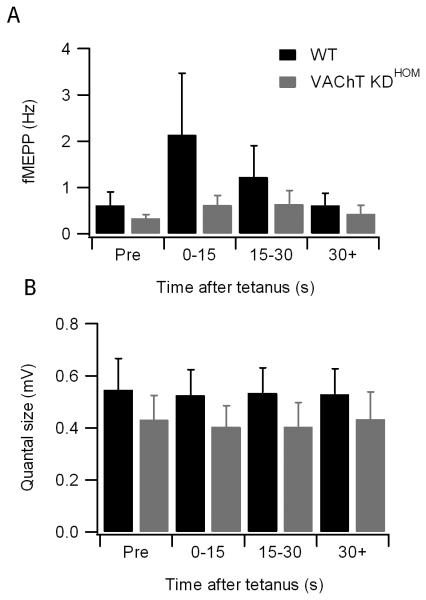
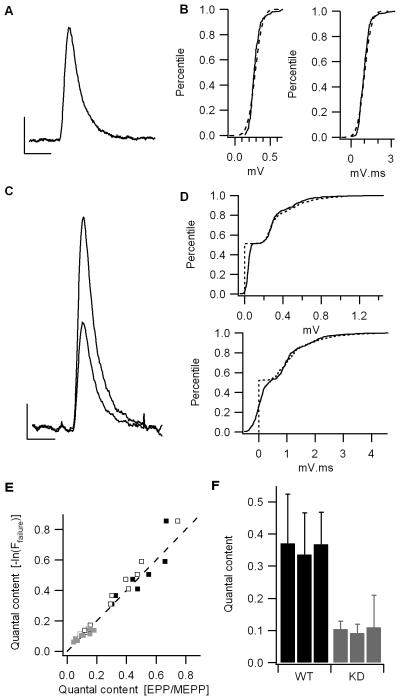
Mammalian motor nerve terminals contain hundreds of thousands of synaptic vesicles, but only a fraction of these vesicles is immediately available for release, the remainder forming a reserve pool. The supply of vesicles is replenished through endocytosis, and newly formed vesicles are refilled with acetylcholine through a process that depends on the vesicular acetylcholine transporter (VAChT). During expression of short-term plasticity, quantal release can be increased, but it is unknown whether this reflects enhanced recruitment of vesicles from the reserve pool or rapid recycling. We examined spontaneous and evoked release of acetylcholine at endplates from genetically modified VAChT KD(HOM) mice that express approximately 30% of the normal level of VAChT to determine steps rate-limited by synaptic vesicle filling. Quantal content and quantal size were reduced in VAChT KD(HOM) mice compared with wild-type controls. Although high-frequency stimulation did not reduce quantal size further, the post-tetanic increase in end-plate potential amplitude or MEPP frequency was significantly smaller in VAChT KD(HOM) mice. This was the case even when tetanic depression was eliminated using an extracellular solution containing reduced Ca(2+) and raised Mg(2+). These results reveal the dependence of short-term plasticity on the level of VAChT expression and efficient synaptic vesicle filling.
Figures
Similar articles
-
Fast and slow-twitching muscles are differentially affected by reduced cholinergic transmission in mice deficient for VAChT: A mouse model for congenital myasthenia.Neurochem Int. 2018 Nov;120:1-12. doi: 10.1016/j.neuint.2018.07.002. Epub 2018 Jul 9. Neurochem Int. 2018. PMID: 30003945 Free PMC article.
-
Reduced expression of the vesicular acetylcholine transporter and neurotransmitter content affects synaptic vesicle distribution and shape in mouse neuromuscular junction.PLoS One. 2013 Nov 8;8(11):e78342. doi: 10.1371/journal.pone.0078342. eCollection 2013. PLoS One. 2013. PMID: 24260111 Free PMC article.
-
The vesicular acetylcholine transporter is required for neuromuscular development and function.Mol Cell Biol. 2009 Oct;29(19):5238-50. doi: 10.1128/MCB.00245-09. Epub 2009 Jul 27. Mol Cell Biol. 2009. PMID: 19635813 Free PMC article.
-
Regulation of cholinergic activity by the vesicular acetylcholine transporter.Biochem J. 2013 Mar 1;450(2):265-74. doi: 10.1042/BJ20121662. Biochem J. 2013. PMID: 23410039 Review.
-
Non-quantal acetylcholine release at the neuromuscular junction.Physiol Res. 2009;58(6):763-784. doi: 10.33549/physiolres.931865. Physiol Res. 2009. PMID: 20059289 Review.
Cited by
-
Vesicular and plasma membrane transporters for neurotransmitters.Cold Spring Harb Perspect Biol. 2012 Feb 1;4(2):a005595. doi: 10.1101/cshperspect.a005595. Cold Spring Harb Perspect Biol. 2012. PMID: 22199021 Free PMC article. Review.
-
Acute Lung Injury in Cholinergic-Deficient Mice Supports Anti-Inflammatory Role of α7 Nicotinic Acetylcholine Receptor.Int J Mol Sci. 2021 Jul 14;22(14):7552. doi: 10.3390/ijms22147552. Int J Mol Sci. 2021. PMID: 34299169 Free PMC article.
-
Drosophila melanogaster as a genetic model system to study neurotransmitter transporters.Neurochem Int. 2014 Jul;73:71-88. doi: 10.1016/j.neuint.2014.03.015. Epub 2014 Apr 3. Neurochem Int. 2014. PMID: 24704795 Free PMC article. Review.
-
Vesicular neurotransmitter transporters in Drosophila melanogaster.Biochim Biophys Acta Biomembr. 2020 Dec 1;1862(12):183308. doi: 10.1016/j.bbamem.2020.183308. Epub 2020 Apr 17. Biochim Biophys Acta Biomembr. 2020. PMID: 32305263 Free PMC article. Review.
-
Fast and slow-twitching muscles are differentially affected by reduced cholinergic transmission in mice deficient for VAChT: A mouse model for congenital myasthenia.Neurochem Int. 2018 Nov;120:1-12. doi: 10.1016/j.neuint.2018.07.002. Epub 2018 Jul 9. Neurochem Int. 2018. PMID: 30003945 Free PMC article.
References
-
- Barstad JA, Lilleheil G. Transversally cut diaphragm preparation from rat. An adjuvant tool in the study of the physiology and pharmacology of the myoneural junction. Arch Int Pharmacodyn Ther. 1968;175:373–390. - PubMed
-
- de Castro BM, Jaeger XD, Martins-Silva C, Lima RDF, Amaral E, Menezes C, Lima P, Neves CML, Pires RG, Gould TW, Welch I, Kushmerick C, Guatimosim C, Izquierdo I, Cammarota M, Rylett RJ, Gomez MV, Caron MG, Oppenheim RW, Prado MAM, Prado VF. The vesicular acetylcholine transporter is required for neuromuscular development and function. Mol Cell Biol. 2009;29:5238–5250. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
