

The impact of intragenic CpG content on gene expression
- PMID: 20203083
- PMCID: PMC2896515
- DOI: 10.1093/nar/gkq115
The impact of intragenic CpG content on gene expression
Abstract
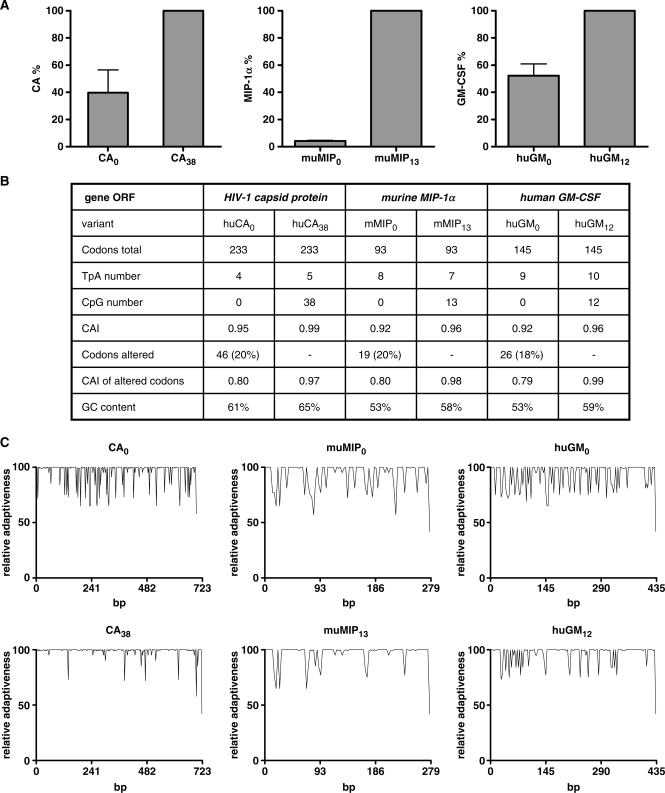
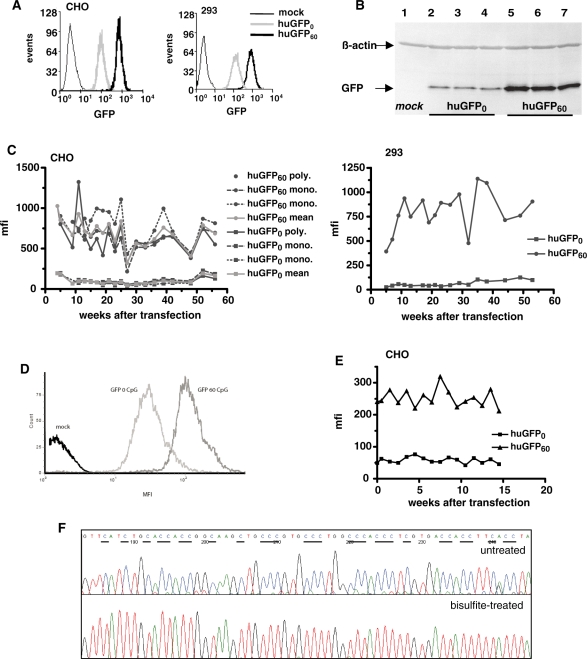
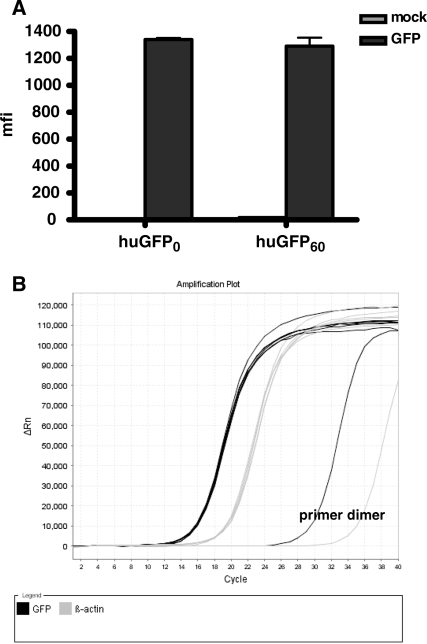
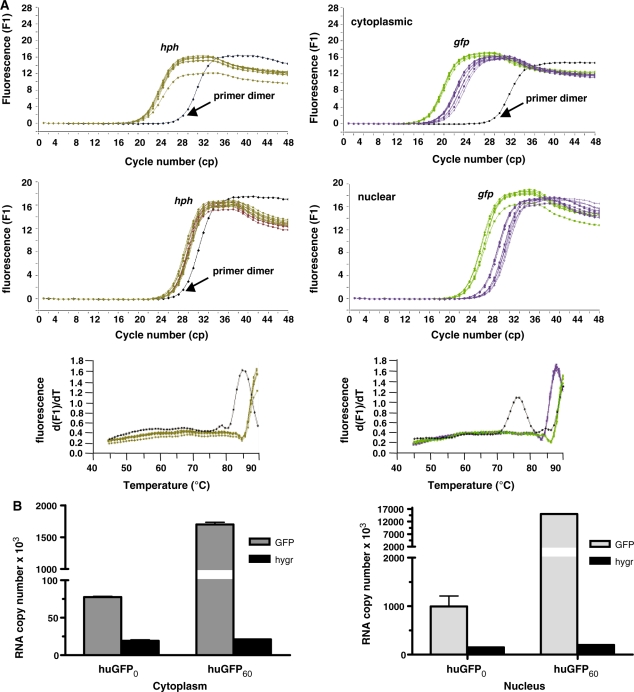
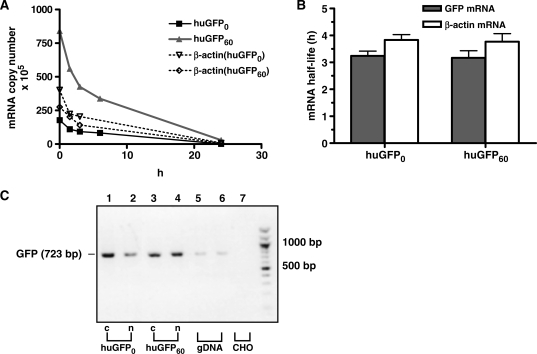
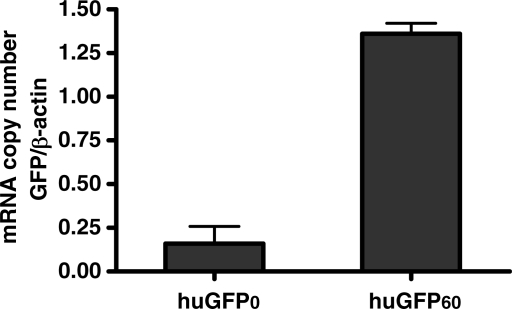
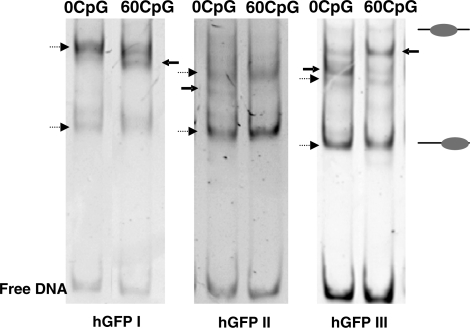
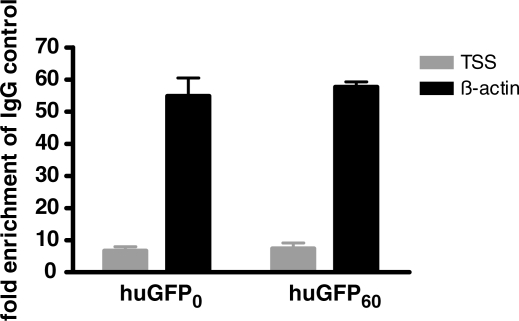
The development of vaccine components or recombinant therapeutics critically depends on sustained expression of the corresponding transgene. This study aimed to determine the contribution of intragenic CpG content to expression efficiency in transiently and stably transfected mammalian cells. Based upon a humanized version of green fluorescent protein (GFP) containing 60 CpGs within its coding sequence, a CpG-depleted variant of the GFP reporter was established by carefully modulating the codon usage. Interestingly, GFP reporter activity and detectable protein amounts in stably transfected CHO and 293 cells were significantly decreased upon CpG depletion and independent from promoter usage (CMV, EF1 alpha). The reduction in protein expression associated with CpG depletion was likewise observed for other unrelated reporter genes and was clearly reflected by a decline in mRNA copy numbers rather than translational efficiency. Moreover, decreased mRNA levels were neither due to nuclear export restrictions nor alternative splicing or mRNA instability. Rather, the intragenic CpG content influenced de novo transcriptional activity thus implying a common transcription-based mechanism of gene regulation via CpGs. Increased high CpG transcription correlated with changed nucleosomal positions in vitro albeit histone density at the two genes did not change in vivo as monitored by ChIP.
Figures


References
-
- Valente CA, Monteiro GA, Cabral JM, Fevereiro M, Prazeres DM. Optimization of the primary recovery of human interferon Alpha2b from Escherichia Coli inclusion bodies. Protein Expr. Purif. 2006;45:226–234. - PubMed
-
- Barnes LM, Dickson AJ. Mammalian cell factories for efficient and stable protein expression. Curr. Opin. Biotechnol. 2006;17:381–386. - PubMed
-
- Jenkins N, Murphy L, Tyther R. Post-Translational modifications of recombinant proteins: significance for biopharmaceuticals. Mol. Biotechnol. 2008;39:113–118. - PubMed
-
- Ill CR, Chiou HC. Gene therapy progress and prospects: recent progress in transgene and Rnai expression cassettes. Gene Ther. 2005;12:795–802. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
