

DNA-PKcs plays a dominant role in the regulation of H2AX phosphorylation in response to DNA damage and cell cycle progression
- PMID: 20205745
- PMCID: PMC2844398
- DOI: 10.1186/1471-2199-11-18
DNA-PKcs plays a dominant role in the regulation of H2AX phosphorylation in response to DNA damage and cell cycle progression
Abstract
Background: When DNA double-strand breaks (DSB) are induced by ionizing radiation (IR) in cells, histone H2AX is quickly phosphorylated into gamma-H2AX (p-S139) around the DSB site. The necessity of DNA-PKcs in regulating the phosphorylation of H2AX in response to DNA damage and cell cycle progression was investigated.
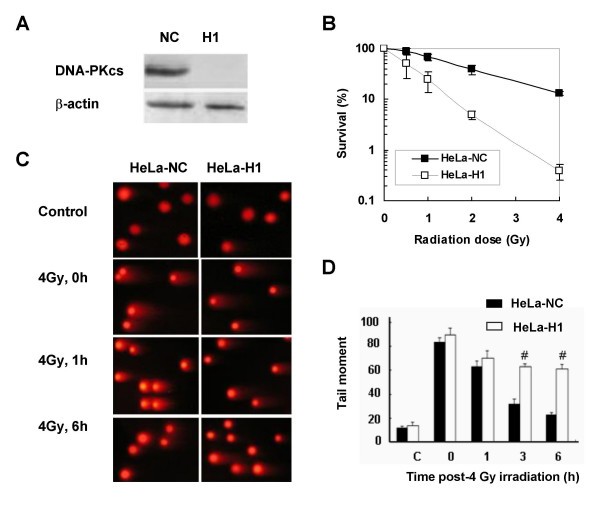
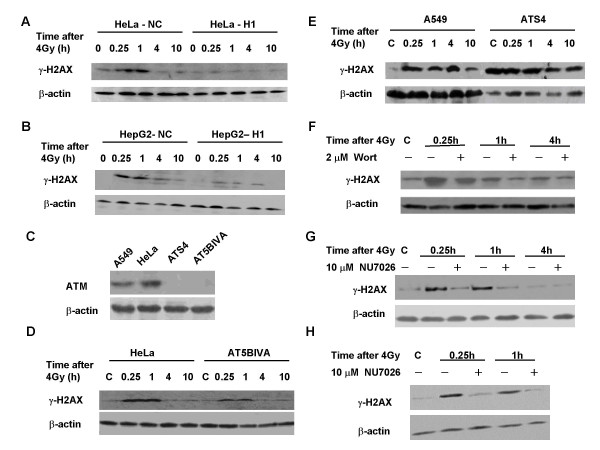
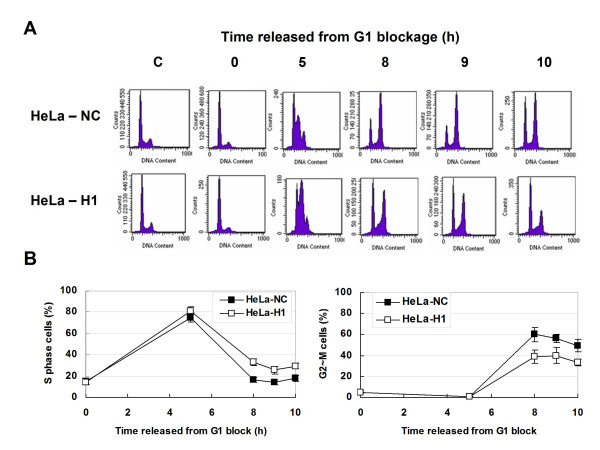
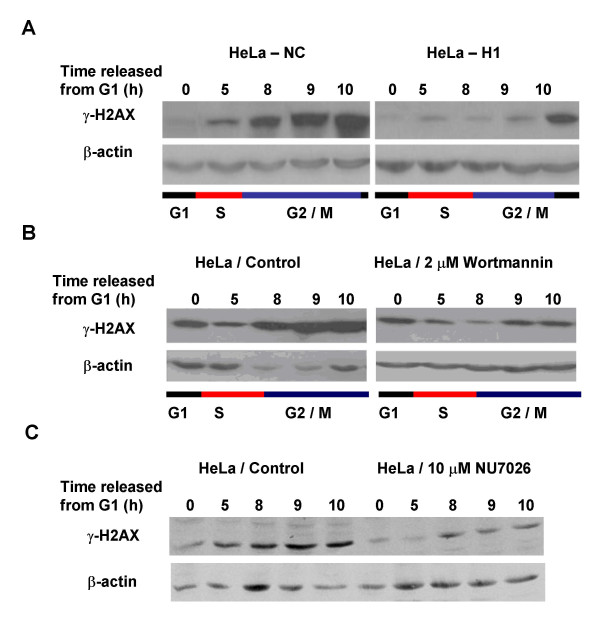
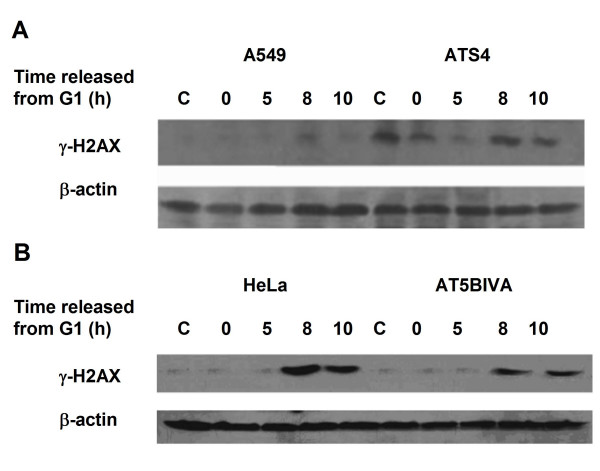
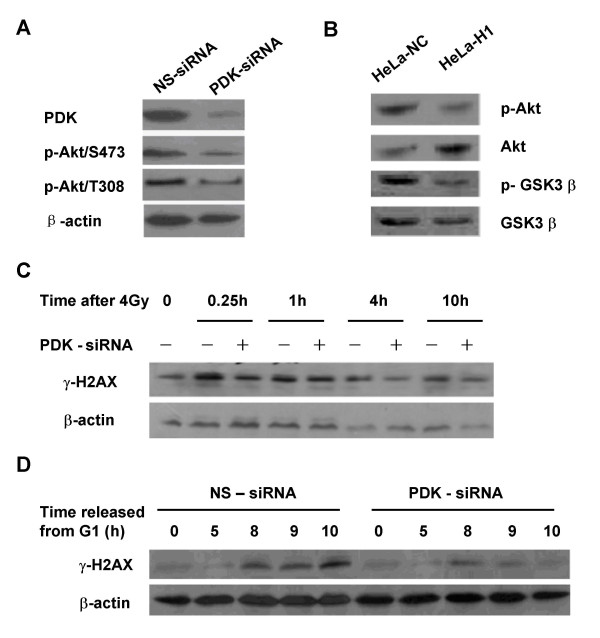
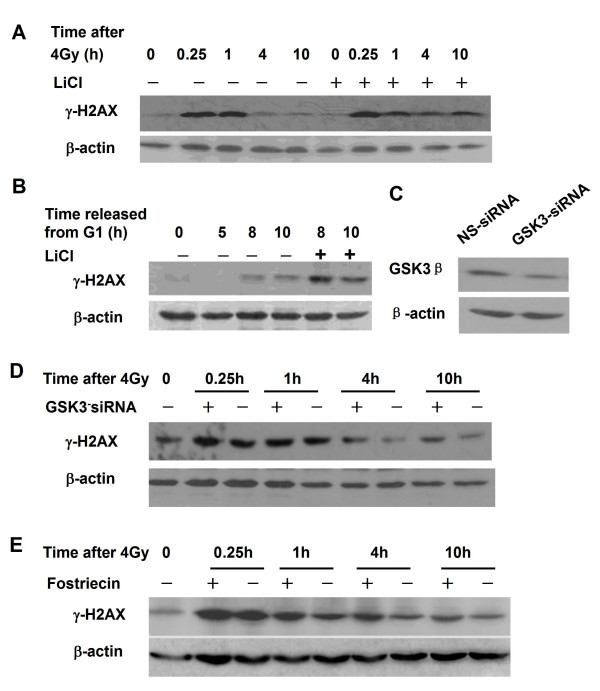
Results: The level of gamma H2AX in HeLa cells increased rapidly with a peak level at 0.25 - 1.0 h after 4 Gy gamma irradiation. SiRNA-mediated depression of DNA-PKcs resulted in a strikingly decreased level of gamma H2AX. An increased gamma H2AX was also induced in the ATM deficient cell line AT5BIVA at 0.5 - 1.0 h after 4 Gy gamma rays, and this IR-increased gamma H2AX in ATM deficient cells was dramatically abolished by the PIKK inhibitor wortmannin and the DNA-PKcs specific inhibitor NU7026. A high level of constitutive expression of gamma H2AX was observed in another ATM deficient cell line ATS4. The alteration of gamma H2AX level associated with cell cycle progression was also observed. HeLa cells with siRNA-depressed DNA-PKcs (HeLa-H1) or normal level DNA-PKcs (HeLa-NC) were synchronized at the G1 phase with the thymidine double-blocking method. At approximately 5 h after the synchronized cells were released from the G1 block, the S phase cells were dominant (80%) for both HeLa-H1 and HeLa-NC cells. At 8 - 9 h after the synchronized cells released from the G1 block, the proportion of G2/M population reached 56 - 60% for HeLa-NC cells, which was higher than that for HeLa H1 cells (33 - 40%). Consistently, the proportion of S phase for HeLa-NC cells decreased to approximately 15%; while a higher level (26 - 33%) was still maintained for the DNA-PKcs depleted HeLa-H1 cells during this period. In HeLa-NC cells, the gamma H2AX level increased gradually as the cells were released from the G1 block and entered the G2/M phase. However, this gamma H2AX alteration associated with cell cycle progressing was remarkably suppressed in the DNA-PKcs depleted HeLa-H1 cells, while wortmannin and NU7026 could also suppress this cell cycle related phosphorylation of H2AX. Furthermore, inhibition of GSK3 beta activity with LiCl or specific siRNA could up-regulate the gamma H2AX level and prolong the time of increased gamma H2AX to 10 h or more after 4 Gy. GSK3 beta is a negative regulation target of DNA-PKcs/Akt signaling via phosphorylation on Ser9, which leads to its inactivation. Depression of DNA-PKcs in HeLa cells leads to a decreased phosphorylation of Akt on Ser473 and its target GSK3 beta on Ser9, which, in other words, results in an increased activation of GSK3 beta. In addition, inhibition of PDK (another up-stream regulator of Akt/GSK3 beta) by siRNA can also decrease the induction of gamma H2AX in response to both DNA damage and cell cycle progression.
Conclusion: DNA-PKcs plays a dominant role in regulating the phosphorylation of H2AX in response to both DNA damage and cell cycle progression. It can directly phosphorylate H2AX independent of ATM and indirectly modulate the phosphorylation level of gamma H2AX via the Akt/GSK3 beta signal pathway.
Figures
References
-
- Albino AP, Huang X, Jorgensen ED, Gietl D, Traganos F, Darzynkiewicz Z. Induction of DNA double-strand breaks in A549 and normal human pulmonary epithelial cells by cigarette smoke is mediated by free radicals. Int J Oncol. 2006;28:1491–505. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
