

Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel enzyme involved in cellular defense against hyperosmotic stress
- PMID: 20207735
- PMCID: PMC2881771
- DOI: 10.1074/jbc.M109.077925
Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel enzyme involved in cellular defense against hyperosmotic stress
Abstract
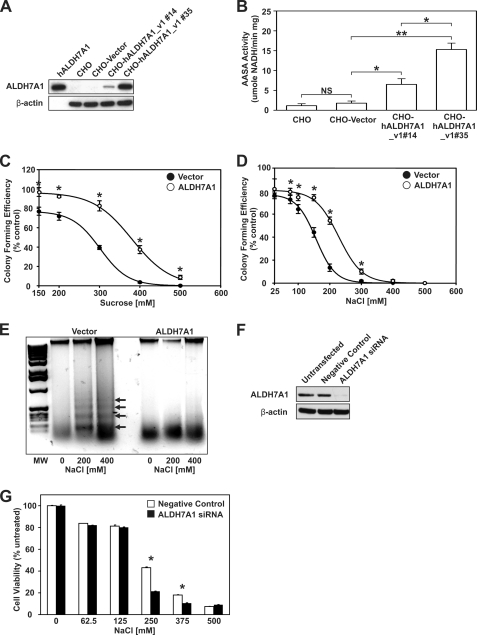
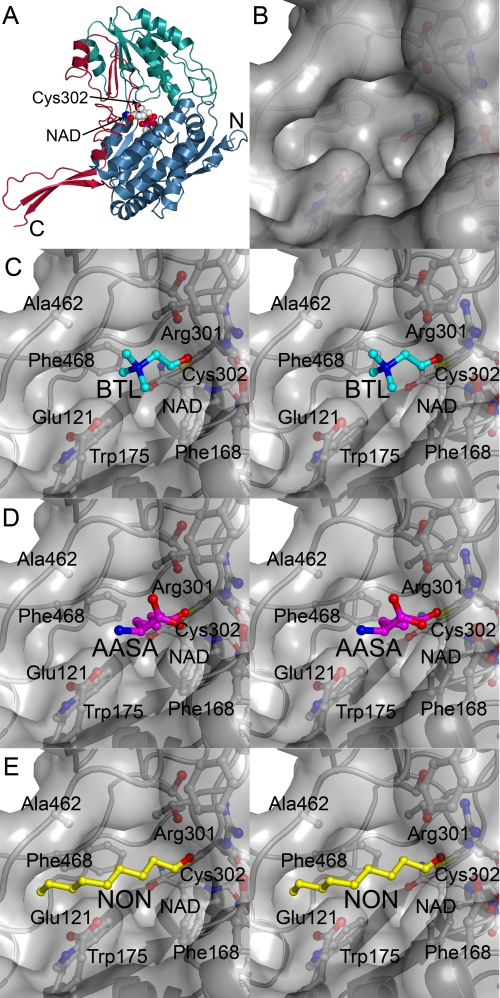
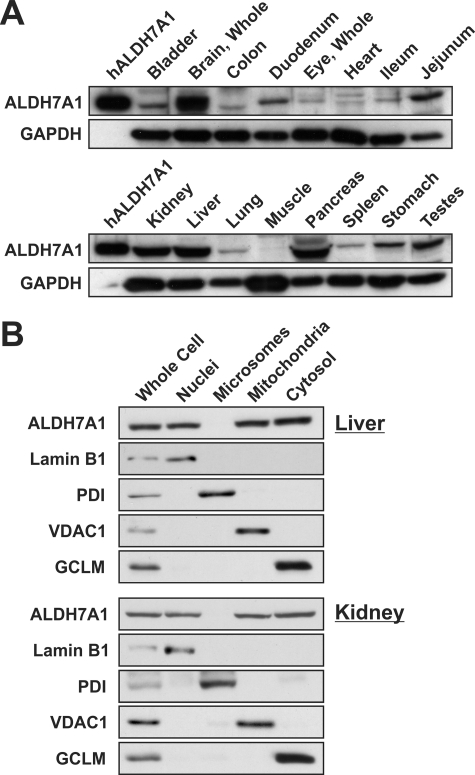
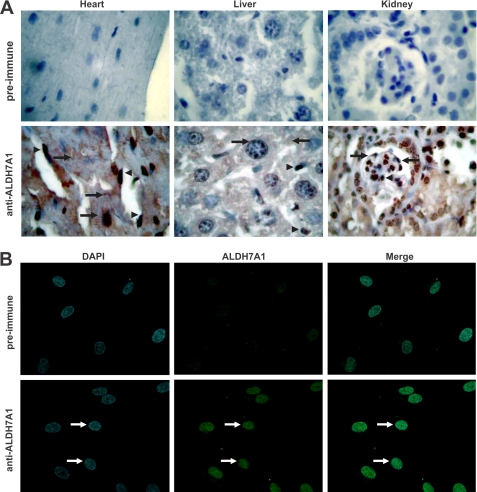
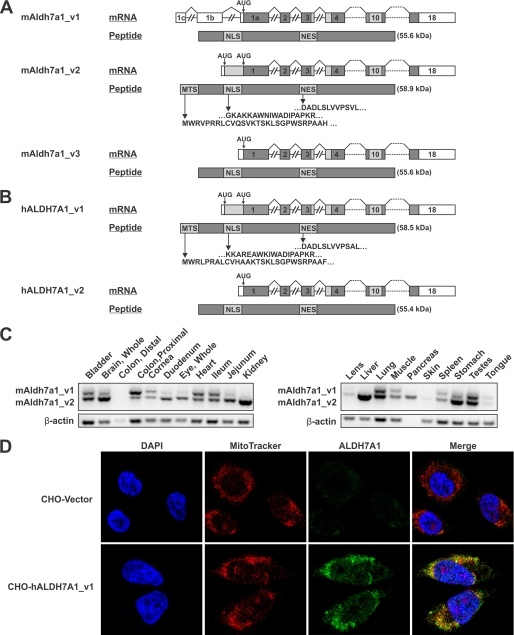
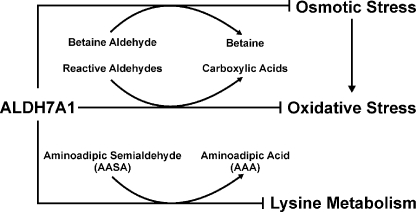
Mammalian ALDH7A1 is homologous to plant ALDH7B1, an enzyme that protects against various forms of stress, such as salinity, dehydration, and osmotic stress. It is known that mutations in the human ALDH7A1 gene cause pyridoxine-dependent and folic acid-responsive seizures. Herein, we show for the first time that human ALDH7A1 protects against hyperosmotic stress by generating osmolytes and metabolizing toxic aldehydes. Human ALDH7A1 expression in Chinese hamster ovary cells attenuated osmotic stress-induced apoptosis caused by increased extracellular concentrations of sucrose or sodium chloride. Purified recombinant ALDH7A1 efficiently metabolized a number of aldehyde substrates, including the osmolyte precursor, betaine aldehyde, lipid peroxidation-derived aldehydes, and the intermediate lysine degradation product, alpha-aminoadipic semialdehyde. The crystal structure for ALDH7A1 supports the enzyme's substrate specificities. Tissue distribution studies in mice showed the highest expression of ALDH7A1 protein in liver, kidney, and brain, followed by pancreas and testes. ALDH7A1 protein was found in the cytosol, nucleus, and mitochondria, making it unique among the aldehyde dehydrogenase enzymes. Analysis of human and mouse cDNA sequences revealed mitochondrial and cytosolic transcripts that are differentially expressed in a tissue-specific manner in mice. In conclusion, ALDH7A1 is a novel aldehyde dehydrogenase expressed in multiple subcellular compartments that protects against hyperosmotic stress by generating osmolytes and metabolizing toxic aldehydes.
Figures
References
-
- Lee P., Kuhl W., Gelbart T., Kamimura T., West C., Beutler E. (1994) Genomics 21, 371–378 - PubMed
-
- Malik H. S., Henikoff S. (2003) Nat. Struct. Biol. 10, 882–891 - PubMed
-
- Guerrero F. D., Jones J. T., Mullet J. E. (1990) Plant Mol. Biol. 15, 11–26 - PubMed
-
- Stroeher V. L., Boothe J. G., Good A. G. (1995) Plant Mol. Biol. 27, 541–551 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
