

Zinc modulates the innate immune response in vivo to polymicrobial sepsis through regulation of NF-kappaB
- PMID: 20207754
- PMCID: PMC2886607
- DOI: 10.1152/ajplung.00368.2009
Zinc modulates the innate immune response in vivo to polymicrobial sepsis through regulation of NF-kappaB
Abstract
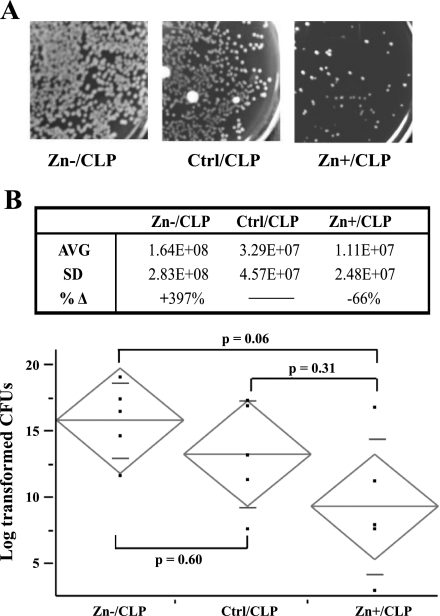
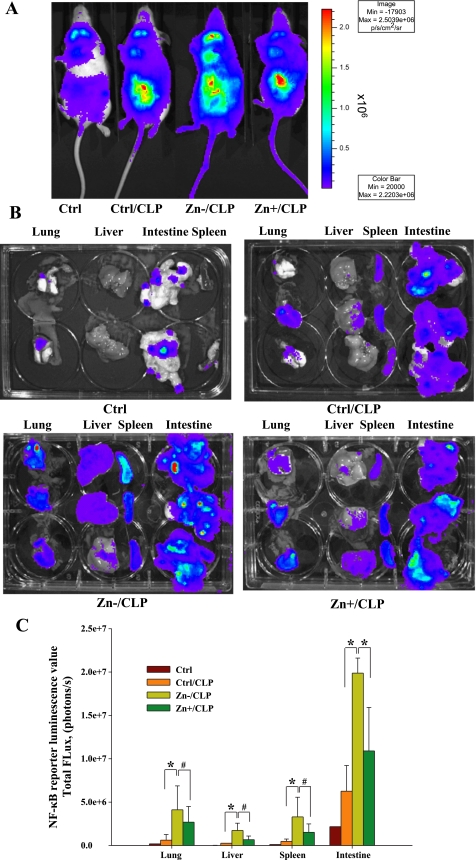
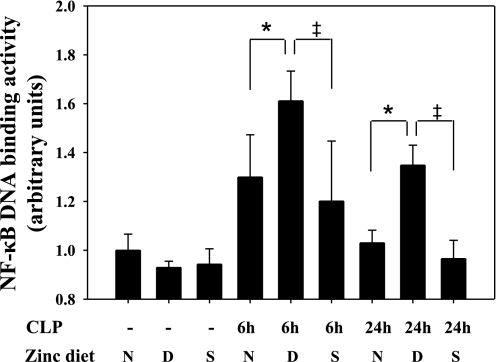
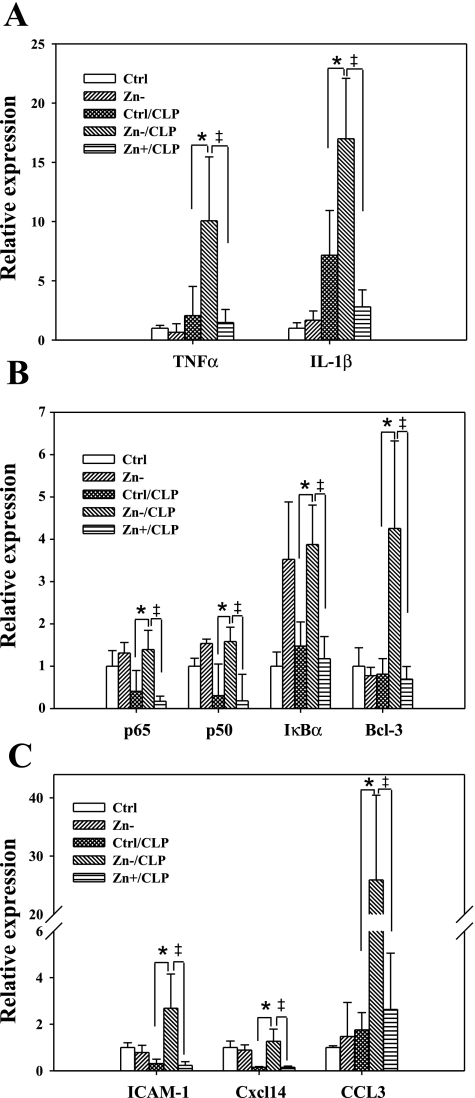
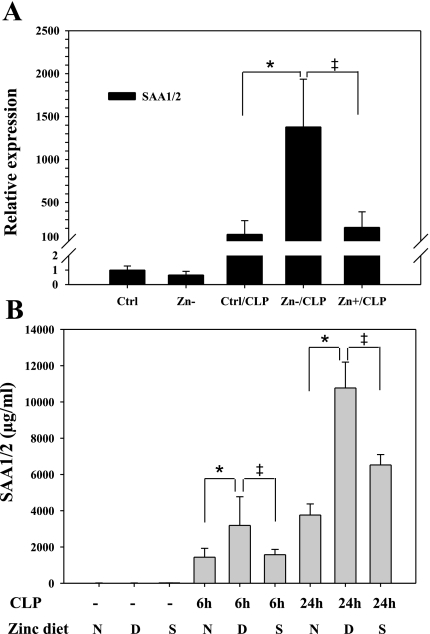
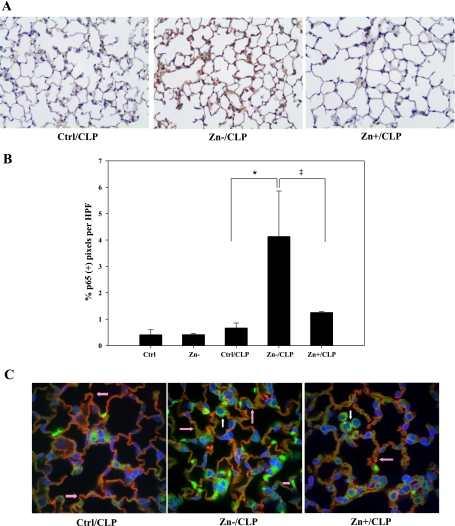
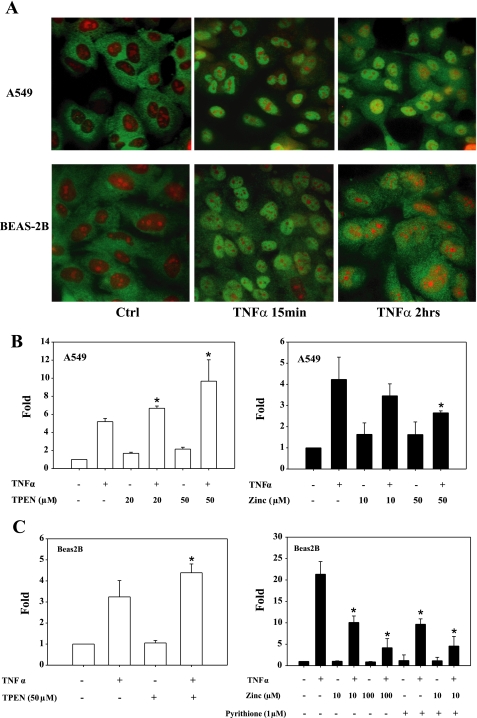
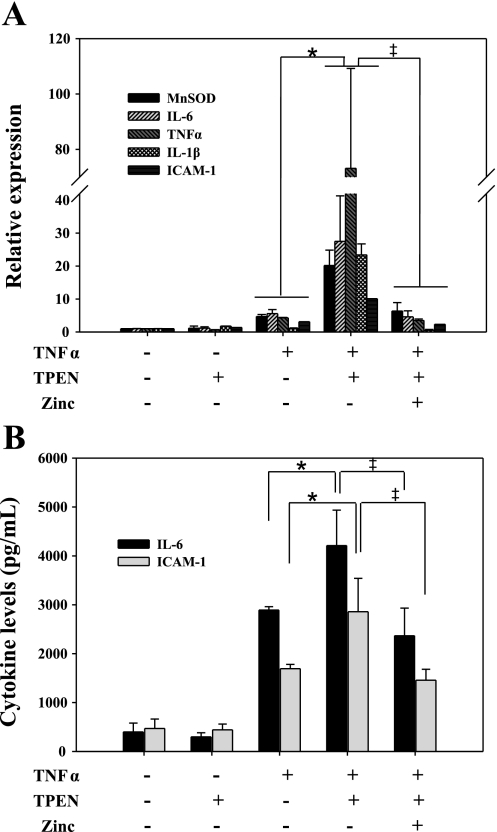
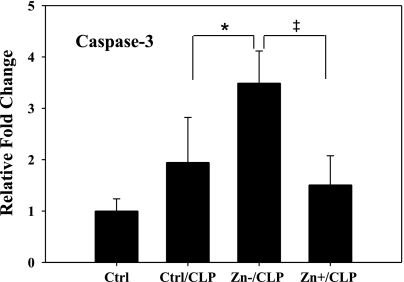
Zinc is an essential element that facilitates coordination of immune activation during the host response to infection. We recently reported that zinc deficiency increases systemic inflammation, vital organ damage, and mortality in a small animal model of sepsis. To investigate potential mechanisms that cause these phenomena, we used the same animal model and observed that zinc deficiency increases bacterial burden and enhances NF-kappaB activity in vital organs including the lung. We conducted further studies in the lung to determine the overall impact of zinc deficiency. At the molecular level, NF-kappaB p65 DNA-binding activity was enhanced by zinc deficiency in response to polymicrobial sepsis. Furthermore, expression of the NF-kappaB-targeted genes IL-1beta, TNFalpha, ICAM-1, and the acute phase response gene SAA1/2 were elevated by zinc deficiency. Unexpectedly, the amount of NF-kappaB p65 mRNA and protein was increased in the lung including alveolar epithelia of zinc-deficient mice. These events occurred with a significant and concomitant increase in caspase-3 activity within 24 h of sepsis onset in zinc-deficient mice relative to control group. Short-term zinc supplementation reversed these effects. Reconstitution of zinc deficiency in lung epithelial cultures resulted in similar findings in response to TNFalpha. Taken together, zinc deficiency systemically enhances the spread of infection and NF-kappaB activation in vivo in response to polymicrobial sepsis, leading to enhanced inflammation, lung injury, and, as reported previously, mortality. Zinc supplementation immediately before initiation of sepsis reversed these effects thereby supporting the plausibility of future studies that explore zinc supplementation strategies to prevent sepsis-mediated morbidity and mortality.
Figures
References
-
- Abraham E, Matthay MA, Dinarello CA, Vincent JL, Cohen J, Opal SM, Glauser M, Parsons P, Fisher CJ, Jr, Repine JE. Consensus conference definitions for sepsis, septic shock, acute lung injury, and acute respiratory distress syndrome: time for a reevaluation. Crit Care Med 28: 232–235, 2000 - PubMed
-
- Abraham E, Singer M. Mechanisms of sepsis-induced organ dysfunction. Crit Care Med 35: 2408–2416, 2007 - PubMed
-
- Balibrea JL, Arias-Diaz J. Acute respiratory distress syndrome in the septic surgical patient. World J Surg 27: 1275–1284, 2003 - PubMed
-
- Bao S, Knoell DL. Zinc modulates airway epithelium susceptibility to death receptor-mediated apoptosis. Am J Physiol Lung Cell Mol Physiol 290: L433–L441, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous