

Alternative end-joining is suppressed by the canonical NHEJ component Xrcc4-ligase IV during chromosomal translocation formation
- PMID: 20208544
- PMCID: PMC3893185
- DOI: 10.1038/nsmb.1773
Alternative end-joining is suppressed by the canonical NHEJ component Xrcc4-ligase IV during chromosomal translocation formation
Abstract
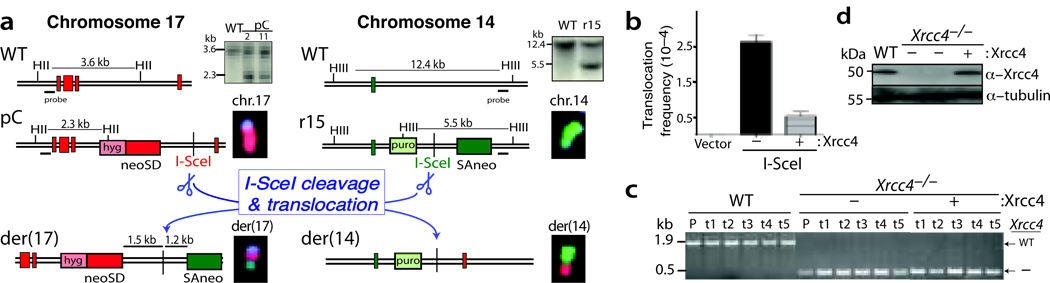
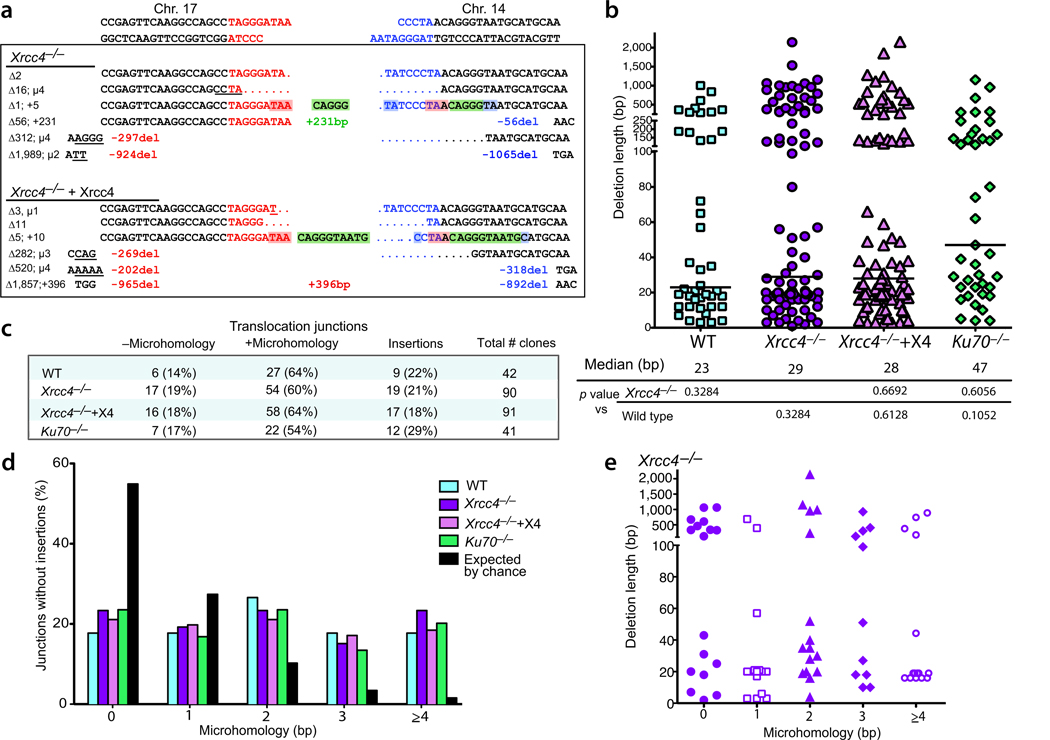
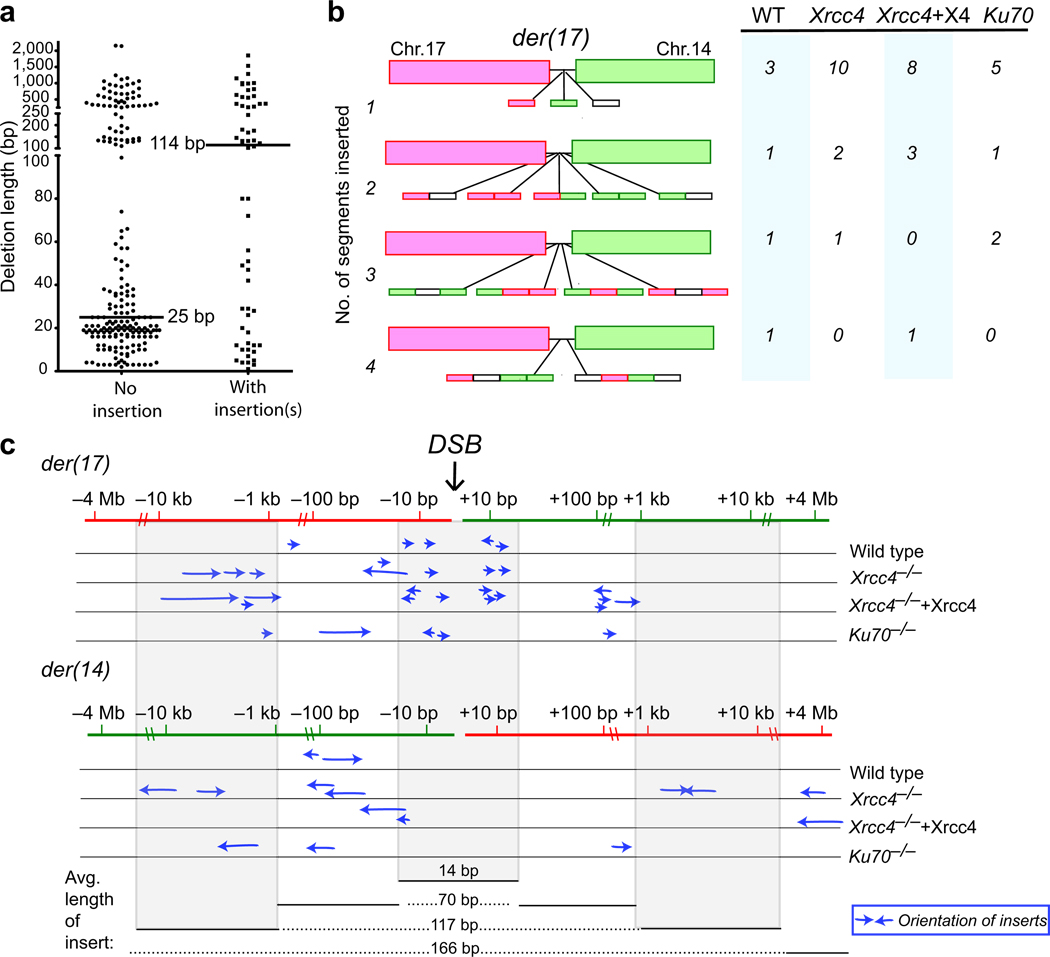
Chromosomal translocations in hematologic and mesenchymal tumors form overwhelmingly by nonhomologous end-joining (NHEJ). Canonical NHEJ, essential for the repair of radiation-induced and some programmed double-strand breaks (DSBs), requires the Xrcc4-ligase IV complex. For other DSBs, the requirement for Xrcc4-ligase IV is less stringent, suggesting the existence of alternative end-joining (alt-NHEJ) pathways. To understand the contributions of the canonical NHEJ and alt-NHEJ pathways, we examined translocation formation in cells deficient in Xrcc4-ligase IV. We found that Xrcc4-ligase IV is not required for but rather suppresses translocations. Translocation breakpoint junctions have similar characteristics in wild-type cells and cells deficient in Xrcc4-ligase IV, including an unchanged bias toward microhomology, unlike what is observed for intrachromosomal DSB repair. Complex insertions in some junctions show that joining can be iterative, encompassing successive processing steps before joining. Our results imply that alt-NHEJ is the primary mediator of translocation formation in mammalian cells.
Figures
Comment in
-
NHEJ and its backup pathways in chromosomal translocations.Nat Struct Mol Biol. 2010 Apr;17(4):393-5. doi: 10.1038/nsmb0410-393. Nat Struct Mol Biol. 2010. PMID: 20368722 Free PMC article. No abstract available.
References
-
- van Gent DC, van der Burg M. Non-homologous end-joining, a sticky affair. Oncogene. 2007;26:7731–7740. - PubMed
-
- Jeggo PA. Identification of genes involved in repair of DNA double-strand breaks in mammalian cells. Radiat Res. 1998;150:S80–S91. - PubMed
-
- Lieber MR, Ma Y, Pannicke U, Schwarz K. Mechanism and regulation of human non-homologous DNA end-joining. Nat Rev Mol Cell Biol. 2003;4:712–720. - PubMed
-
- Chaudhuri J, et al. Evolution of the immunoglobulin heavy chain class switch recombination mechanism. Adv Immunol. 2007;94:157–214. - PubMed
-
- Greaves MF, Wiemels J. Origins of chromosome translocations in childhood leukaemia. Nat Rev Cancer. 2003;3:639–649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
