

Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of Dicer-like proteins
- PMID: 20209086
- PMCID: PMC2831073
- DOI: 10.1371/journal.pone.0009514
Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of Dicer-like proteins
Erratum in
- PLoS One. 2010;5(4). doi: 10.1371/annotation/726f31b5-99c4-44e9-9cd6-b8d66b3f6038. Hollander, Jens [corrected to Hollunder, Jens] doi: 10.1371/annotation/726f31b5-99c4-44e9-9cd6-b8d66b3f6038
Abstract
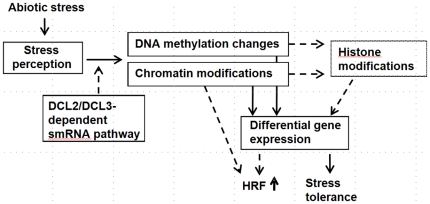
Epigenetic states and certain environmental responses in mammals and seed plants can persist in the next sexual generation. These transgenerational effects have potential adaptative significance as well as medical and agronomic ramifications. Recent evidence suggests that some abiotic and biotic stress responses of plants are transgenerational. For example, viral infection of tobacco plants and exposure of Arabidopsis thaliana plants to UVC and flagellin can induce transgenerational increases in homologous recombination frequency (HRF). Here we show that exposure of Arabidopsis plants to stresses, including salt, UVC, cold, heat and flood, resulted in a higher HRF, increased global genome methylation, and higher tolerance to stress in the untreated progeny. This transgenerational effect did not, however, persist in successive generations. Treatment of the progeny of stressed plants with 5-azacytidine was shown to decrease global genomic methylation and enhance stress tolerance. Dicer-like (DCL) 2 and DCL3 encode Dicer activities important for small RNA-dependent gene silencing. Stress-induced HRF and DNA methylation were impaired in dcl2 and dcl3 deficiency mutants, while in dcl2 mutants, only stress-induced stress tolerance was impaired. Our results are consistent with the hypothesis that stress-induced transgenerational responses in Arabidopsis depend on altered DNA methylation and smRNA silencing pathways.
Conflict of interest statement
Figures
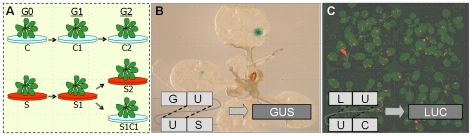
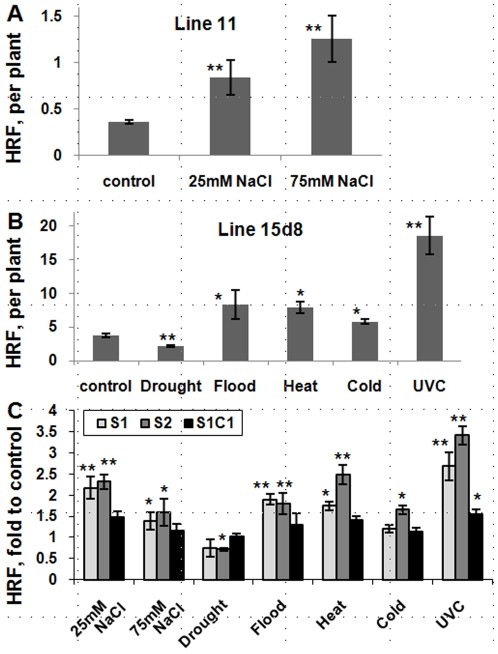
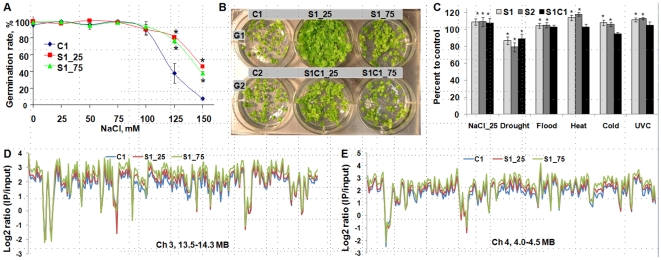
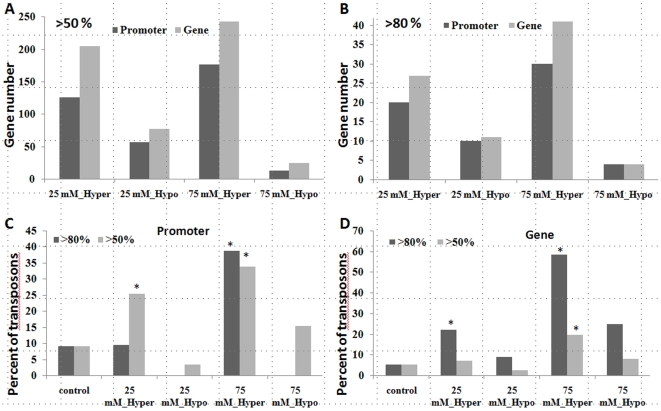
References
-
- Koturbash I, Baker M, Loree J, Kutanzi K, Hudson D, et al. Epigenetic dysregulation underlies radiation-induced transgenerational genome instability in vivo. Int J Radiat Oncol Biol Phys. 2006;66:327–330. - PubMed
-
- Molinier J, Ries G, Zipfel C, Hohn B. Transgeneration memory of stress in plants. Nature. 2006;442:1046–1049. - PubMed
-
- Pembrey ME, Bygren LO, Kaati G, Edvinsson S, Northstone K, et al. Sex-specific, male-line transgenerational responses in humans. Eur J Hum Genet. 2006;14:159–166. - PubMed
-
- Blodner C, Goebel C, Feussner I, Gatz C, Polle A. Warm and cold parental reproductive environments affect seed properties, fitness, and cold responsiveness in Arabidopsis thaliana progenies. Plant Cell Environ. 2007;30:165–175. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
