

A paucity of heterochromatin at functional human neocentromeres
- PMID: 20210998
- PMCID: PMC2845132
- DOI: 10.1186/1756-8935-3-6
A paucity of heterochromatin at functional human neocentromeres
Abstract
Background: Centromeres are responsible for the proper segregation of replicated chromatids during cell division. Neocentromeres are fully functional ectopic human centromeres that form on low-copy DNA sequences and permit analysis of centromere structure in relation to the underlying DNA sequence. Such structural analysis is not possible at endogenous centromeres because of the large amounts of repetitive alpha satellite DNA present.
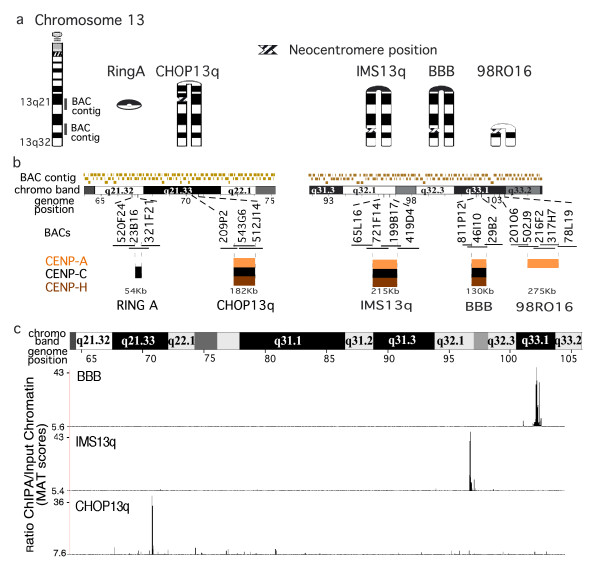
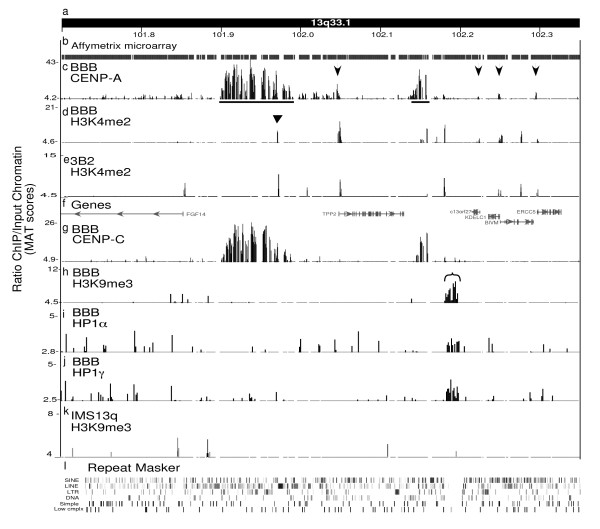
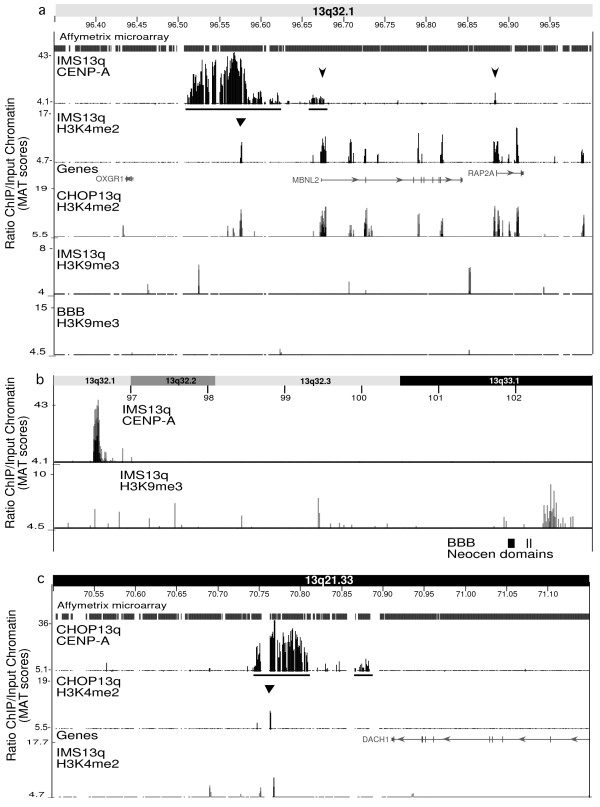
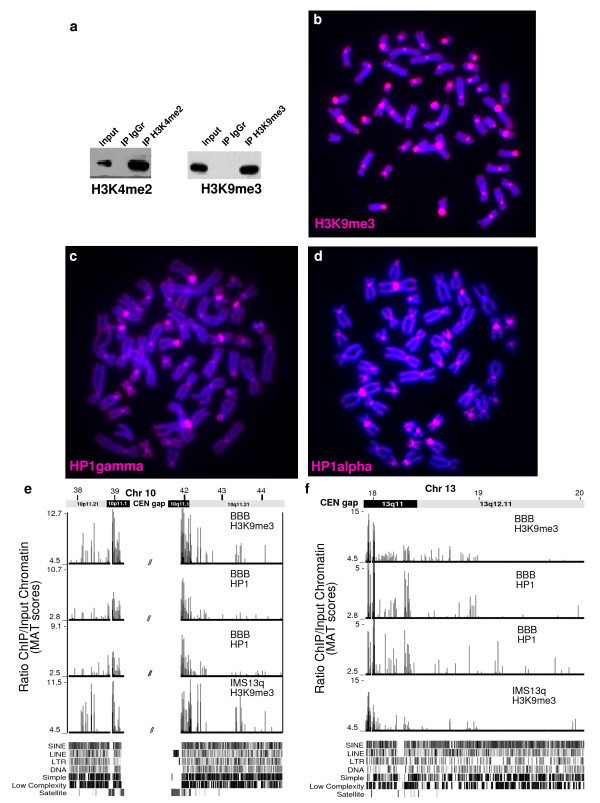
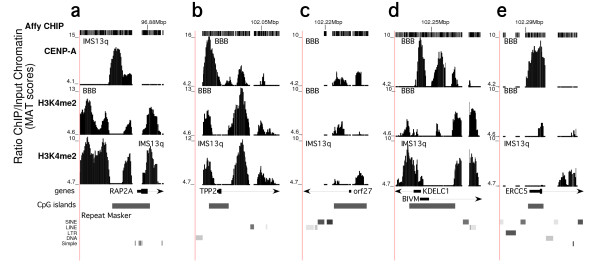
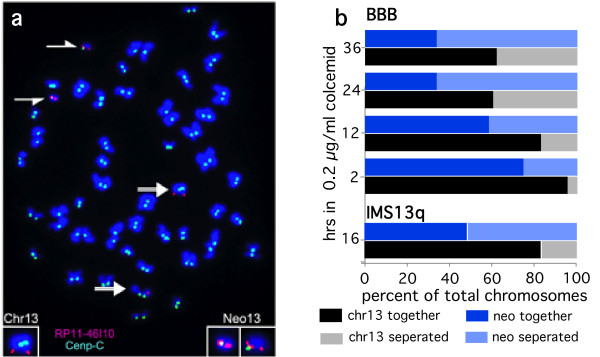
Results: High-resolution chromatin immunoprecipitation (ChIP) on CHIP (microarray) analysis of three independent neocentromeres from chromosome 13q revealed that each neocentromere contained approximately 100 kb of centromere protein (CENP)-A in a two-domain organization. Additional CENP-A domains were observed in the vicinity of neocentromeres, coinciding with CpG islands at the 5' end of genes. Analysis of histone H3 dimethylated at lysine 4 (H3K4me2) revealed small domains at each neocentromere. However, these domains of H3K4me2 were also found in the equivalent non-neocentric chromosomes. A surprisingly minimal (approximately 15 kb) heterochromatin domain was observed at one of the neocentromeres, which formed in an unusual transposon-free region distal to the CENP-A domains. Another neocentromere showed a distinct absence of nearby significant domains of heterochromatin. A subtle defect in centromere cohesion detected at these neocentromeres may be due to the paucity of heterochromatin domains.
Conclusions: This high-resolution mapping suggests that H3K4me2 does not seem sufficiently abundant to play a structural role at neocentromeres, as proposed for endogenous centromeres. Large domains of heterochromatin also do not appear necessary for centromere function. Thus, this study provides important insight into the structural requirements of human centromere function.
Figures
References
LinkOut - more resources
Full Text Sources
