

Reduction of beta-amyloid pathology by celastrol in a transgenic mouse model of Alzheimer's disease
- PMID: 20211007
- PMCID: PMC2841120
- DOI: 10.1186/1742-2094-7-17
Reduction of beta-amyloid pathology by celastrol in a transgenic mouse model of Alzheimer's disease
Abstract
Background: Abeta deposits represent a neuropathological hallmark of Alzheimer's disease (AD). Both soluble and insoluble Abeta species are considered to be responsible for initiating the pathological cascade that eventually leads to AD. Therefore, the identification of therapeutic approaches that can lower Abeta production or accumulation remains a priority. NFkappaB has been shown to regulate BACE-1 expression level, the rate limiting enzyme responsible for the production of Abeta. We therefore explored whether the known NFkappaB inhibitor celastrol could represent a suitable compound for decreasing Abeta production and accumulation in vivo.
Methods: The effect of celastrol on amyloid precursor protein (APP) processing, Abeta production and NFkappaB activation was investigated by western blotting and ELISAs using a cell line overexpressing APP. The impact of celastrol on brain Abeta accumulation was tested in a transgenic mouse model of AD overexpressing the human APP695sw mutation and the presenilin-1 mutation M146L (Tg PS1/APPsw) by immunostaining and ELISAs. An acute treatment with celastrol was investigated by administering celastrol intraperitoneally at a dosage of 1 mg/Kg in 35 week-old Tg PS1/APPsw for 4 consecutive days. In addition, a chronic treatment (32 days) with celastrol was tested using a matrix-driven delivery pellet system implanted subcutaneously in 5 month-old Tg PS1/APPsw to ensure a continuous daily release of 2.5 mg/Kg of celastrol.
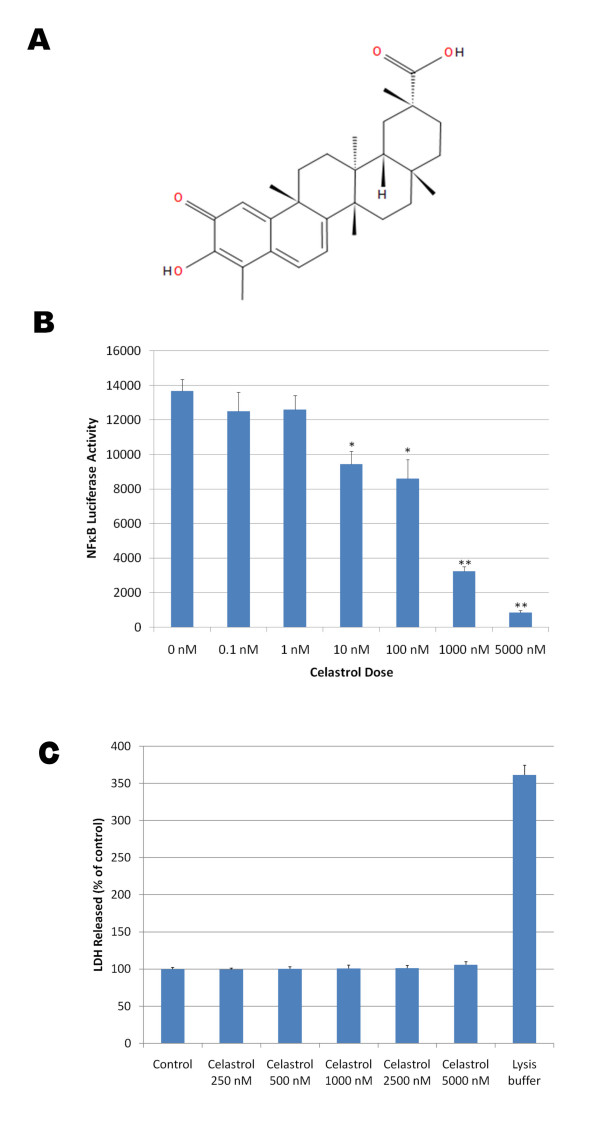
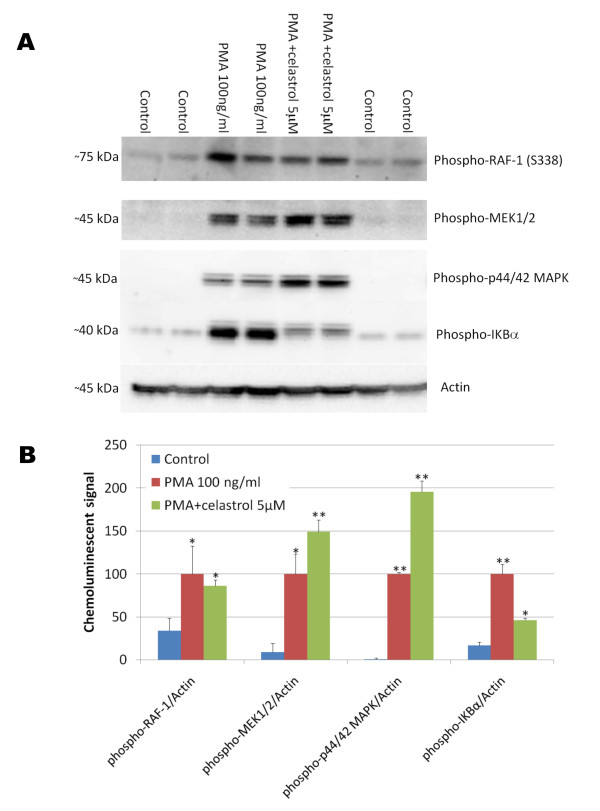
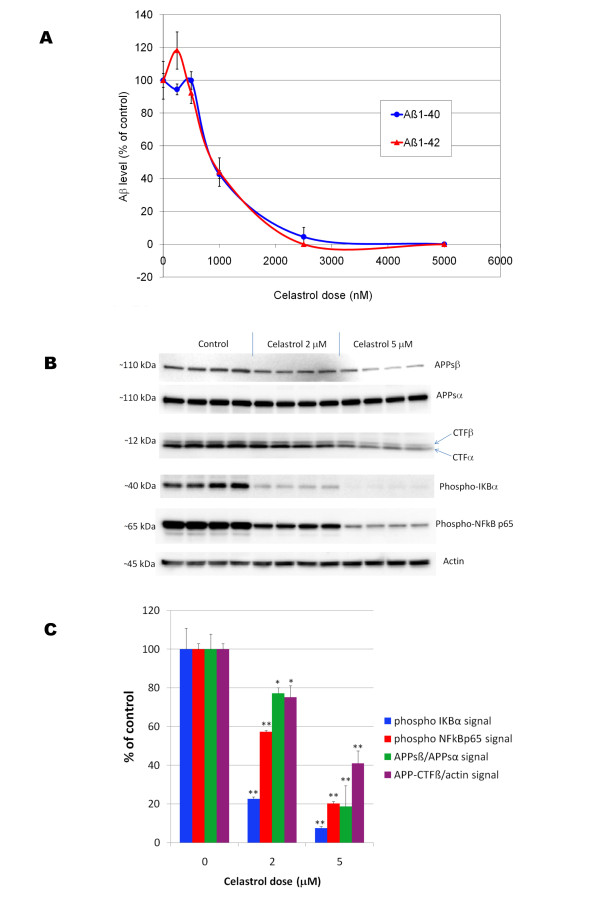
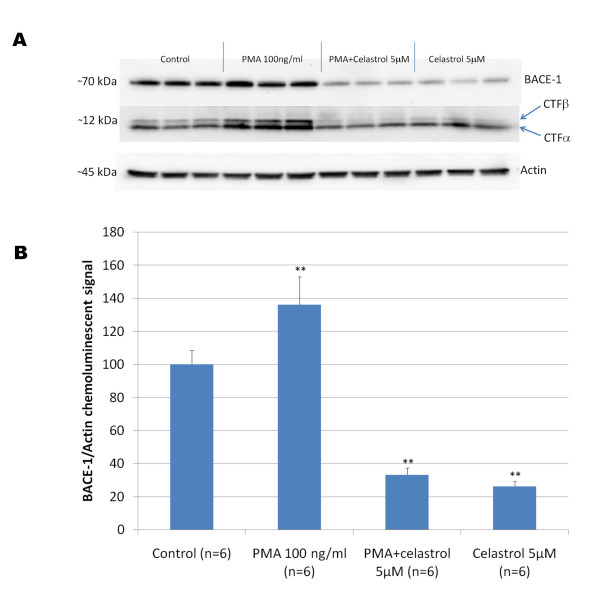
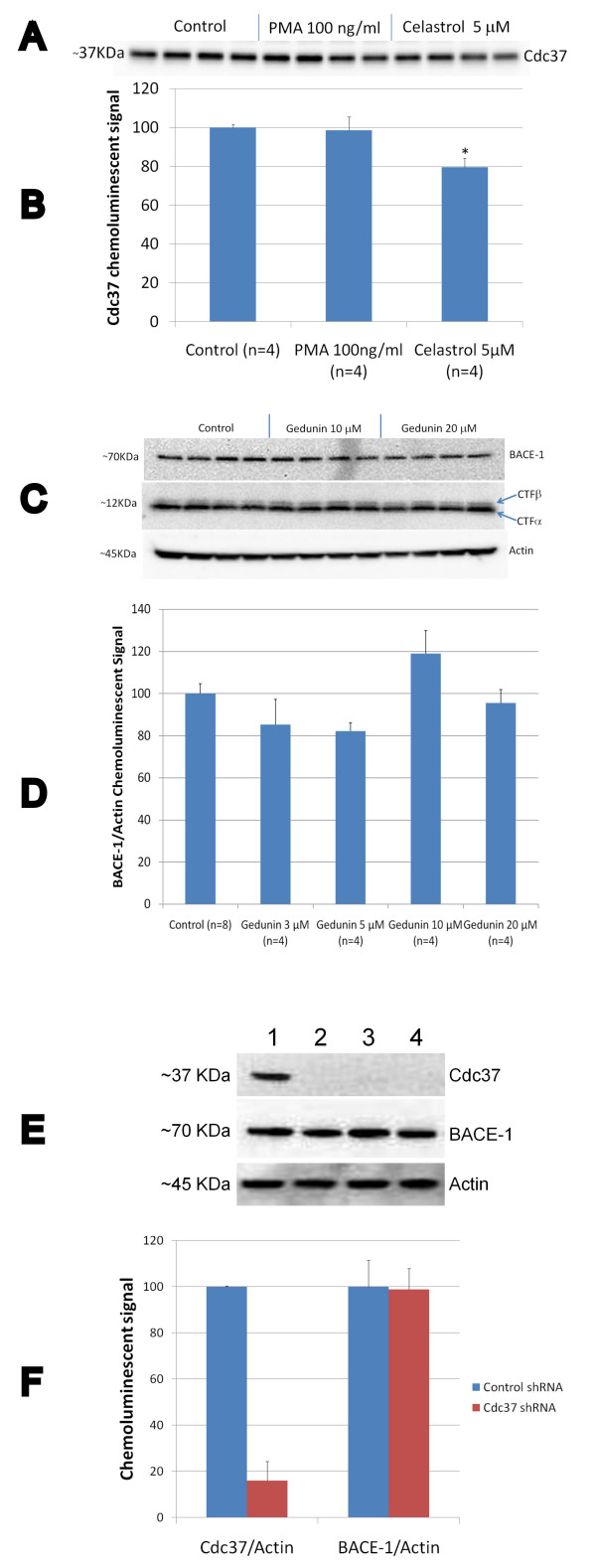
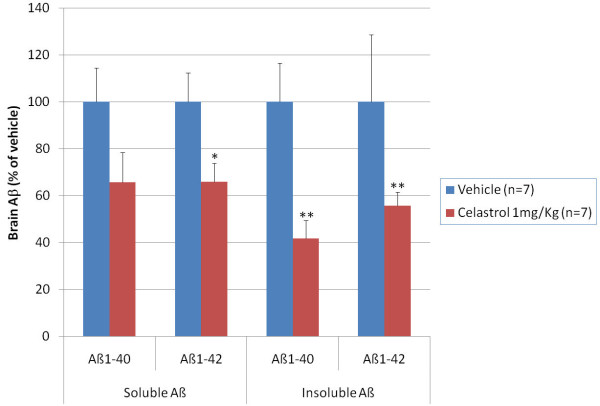
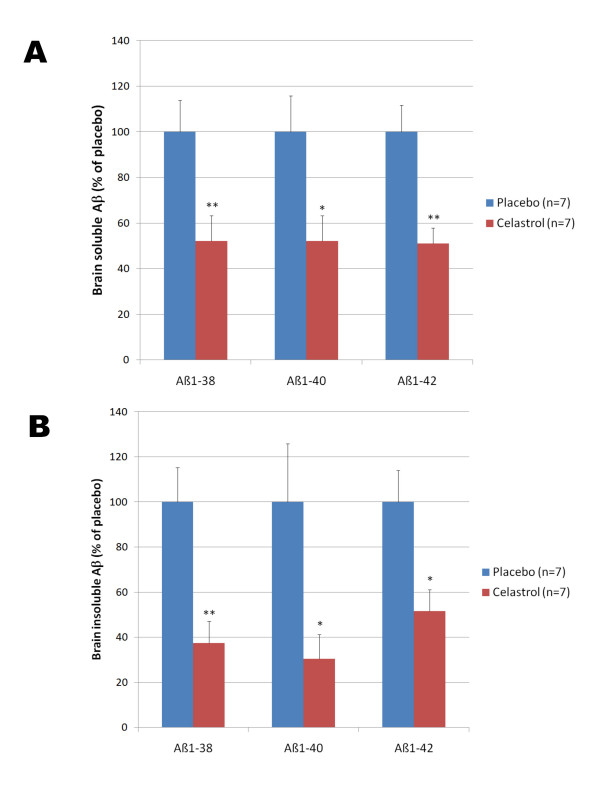
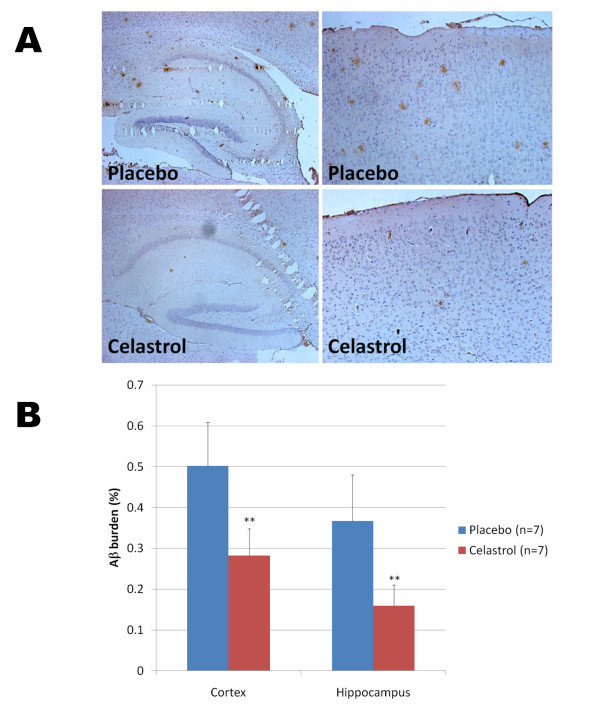
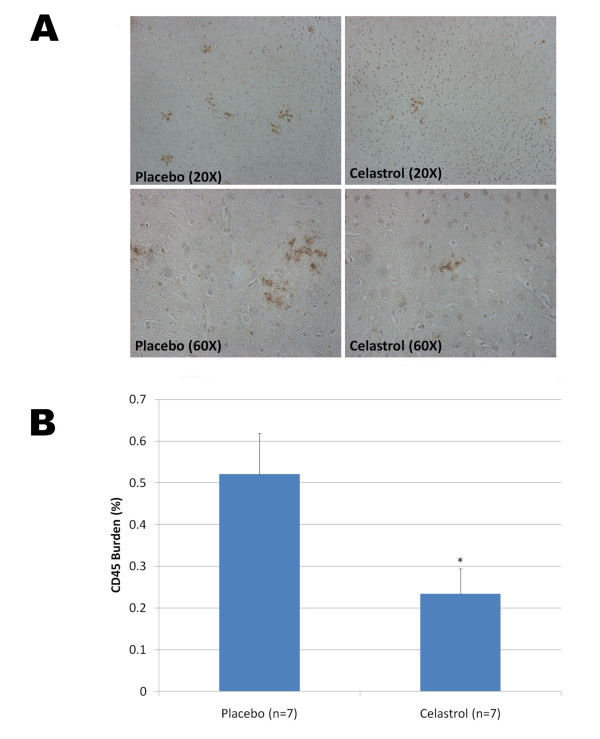
Results: In vitro, celastrol dose dependently prevented NFkappaB activation and inhibited BACE-1 expression. Celastrol potently inhibited Abeta1-40 and Abeta1-42 production by reducing the beta-cleavage of APP, leading to decreased levels of APP-CTFbeta and APPsbeta. In vivo, celastrol appeared to reduce the levels of both soluble and insoluble Abeta1-38, Abeta1-40 and Abeta1-42. In addition, a reduction in Abeta plaque burden and microglial activation was observed in the brains of Tg PS1/APPsw following a chronic administration of celastrol.
Conclusions: Overall our data suggest that celastrol is a potent Abeta lowering compound that acts as an indirect BACE-1 inhibitor possibly by regulating BACE-1 expression level via an NFkappaB dependent mechanism. Additional work is required to determine whether chronic administration of celastrol can be safely achieved with cognitive benefits in a transgenic mouse model of AD.
Figures
References
-
- Vassar R, Bennett BD, Babu-Khan S, Kahn S, Mendiaz EA, Denis P, Teplow DB, Ross S, Amarante P, Loeloff R, Luo Y, Fisher S, Fuller J, Edenson S, Lile J, Jarosinski MA, Biere AL, Curran E, Burgess T, Louis JC, Collins F, Treanor J, Rogers G, Citron M. Beta-secretase cleavage of Alzheimer's amyloid precursor protein by the transmembrane aspartic protease BACE. Science. 1999;286(5440):735–41. doi: 10.1126/science.286.5440.735. - DOI - PubMed
-
- Sinha S, Anderson JP, Barbour R, Basi GS, Caccavello R, Davis D, Doan M, Dovey HF, Frigon N, Hong J, Jacobson-Croak K, Jewett N, Keim P, Knops J, Lieberburg I, Power M, Tan H, Tatsuno G, Tung J, Schenk D, Seubert P, Suomensaari SM, Wang S, Walker D, Zhao J, McConlogue L, John V. Purification and cloning of amyloid precursor protein beta-secretase from human brain. Nature. 1999;402(6761):537–40. doi: 10.1038/990114. - DOI - PubMed
-
- Yan R, Bienkowski MJ, Shuck ME, Miao H, Tory MC, Pauley AM, Brashier JR, Stratman NC, Mathews WR, Buhl AE, Carter DB, Tomasselli AG, Parodi LA, Heinrikson RL, Gurney ME. Membrane-anchored aspartyl protease with Alzheimer's disease beta-secretase activity. Nature. 1999;402(6761):533–7. doi: 10.1038/990107. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
