

New insights into the evolution of subtilisin-like serine protease genes in Pezizomycotina
- PMID: 20211028
- PMCID: PMC2848655
- DOI: 10.1186/1471-2148-10-68
New insights into the evolution of subtilisin-like serine protease genes in Pezizomycotina
Abstract
Background: Subtilisin-like serine proteases play an important role in pathogenic fungi during the penetration and colonization of their hosts. In this study, we perform an evolutionary analysis of the subtilisin-like serine protease genes of subphylum Pezizomycotina to find if there are similar pathogenic mechanisms among the pathogenic fungi with different life styles, which utilize subtilisin-like serine proteases as virulence factors. Within Pezizomycotina, nematode-trapping fungi are unique because they capture soil nematodes using specialized trapping devices. Increasing evidence suggests subtilisin-like serine proteases from nematode-trapping fungi are involved in the penetration and digestion of nematode cuticles. Here we also conduct positive selection analysis on the subtilisin-like serine protease genes from nematode-trapping fungi.
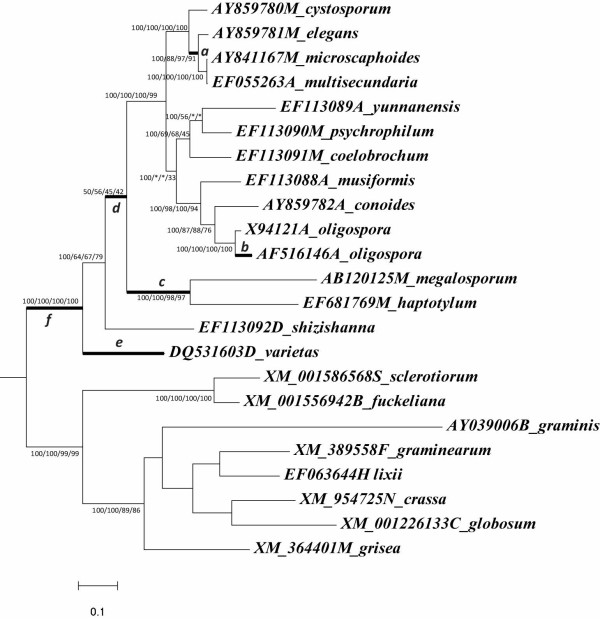
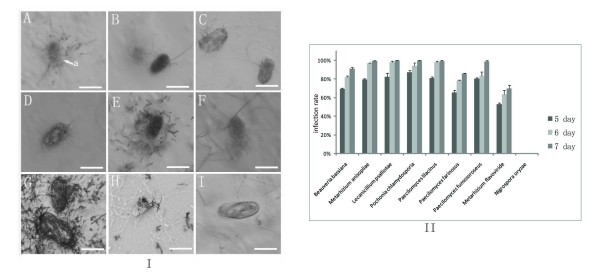
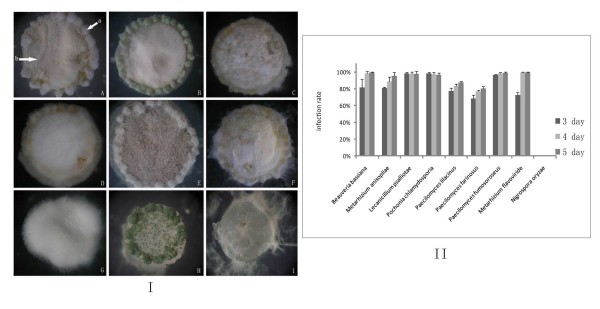
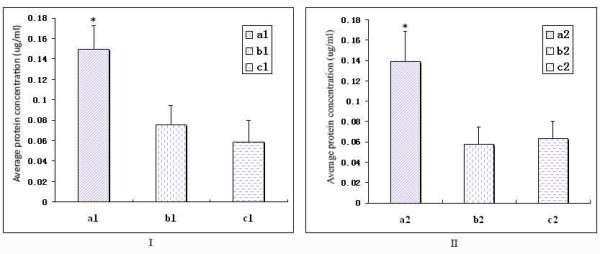
Results: Phylogenetic analysis of 189 subtilisin-like serine protease genes from Pezizomycotina suggests five strongly-supported monophyletic clades. The subtilisin-like serine protease genes previously identified or presumed as endocellular proteases were clustered into one clade and diverged the earliest in the phylogeny. In addition, the cuticle-degrading protease genes from entomopathogenic and nematode-parasitic fungi were clustered together, indicating that they might have overlapping pathogenic mechanisms against insects and nematodes. Our experimental bioassays supported this conclusion. Interestingly, although they both function as cuticle-degrading proteases, the subtilisin-like serine protease genes from nematode-trapping fungi and nematode-parasitic fungi were not grouped together in the phylogenetic tree. Our evolutionary analysis revealed evidence for positive selection on the subtilisin-like serine protease genes of the nematode-trapping fungi.
Conclusions: Our study provides new insights into the evolution of subtilisin-like serine protease genes in Pezizomycotina. Pezizomycotina subtilisins most likely evolved from endocellular to extracellular proteases. The entomopathogenic and nematode-parasitic fungi likely share similar properties in parasitism. In addition, our data provided better understanding about the duplications and subsequent functional divergence of subtilisin-like serine protease genes in Pezizomycotina. The evidence of positive selection detected in the subtilisin-like serine protease genes of nematode-trapping fungi in the present study suggests that the subtilisin-like serine proteases may have played important roles during the evolution of pathogenicity of nematode-trapping fungi against nematodes.
Figures


References
-
- St Leger RJ. The role of cuticle-degrading proteases in fungal pathogenesis of insects. Can J Bot. 1995;73:1119–1125. doi: 10.1139/b95-367. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
