

microRNA-29a induces aberrant self-renewal capacity in hematopoietic progenitors, biased myeloid development, and acute myeloid leukemia
- PMID: 20212066
- PMCID: PMC2839143
- DOI: 10.1084/jem.20090831
microRNA-29a induces aberrant self-renewal capacity in hematopoietic progenitors, biased myeloid development, and acute myeloid leukemia
Abstract
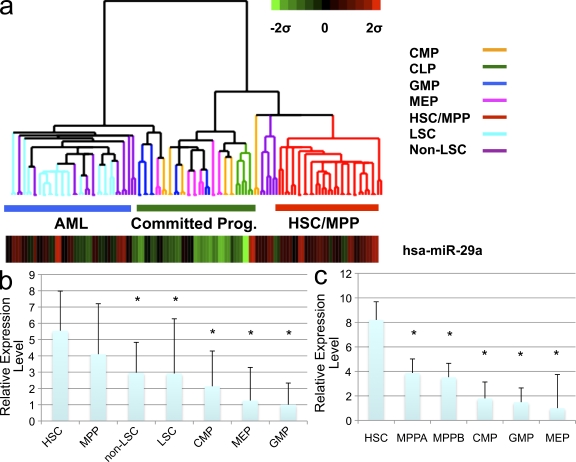
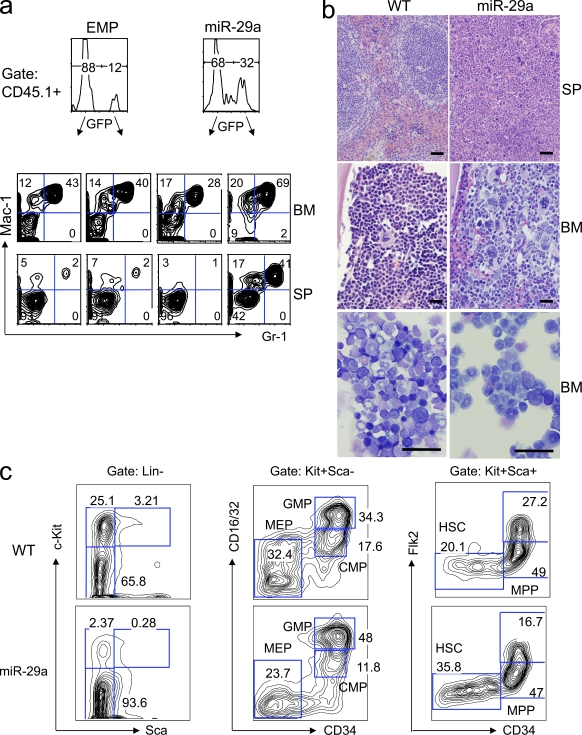
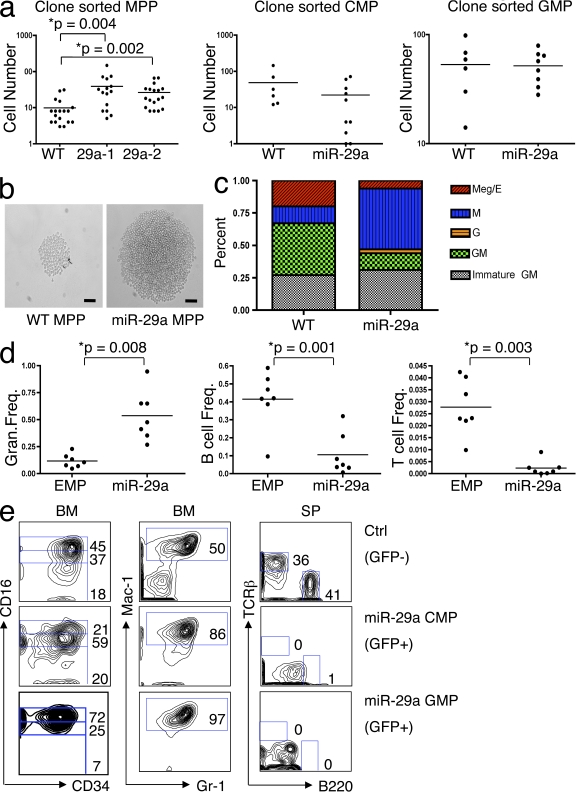
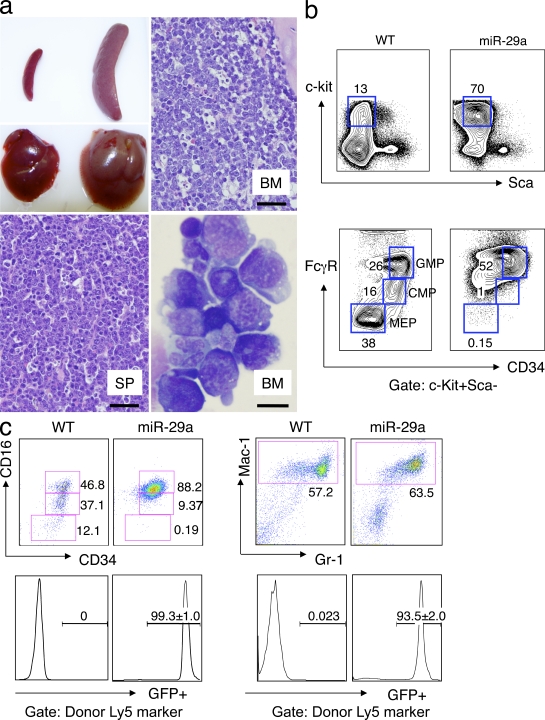
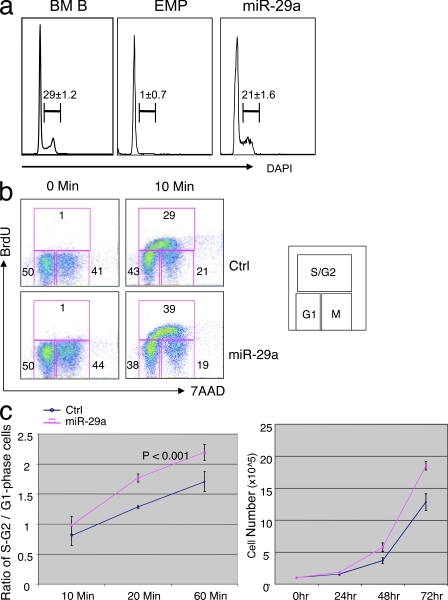
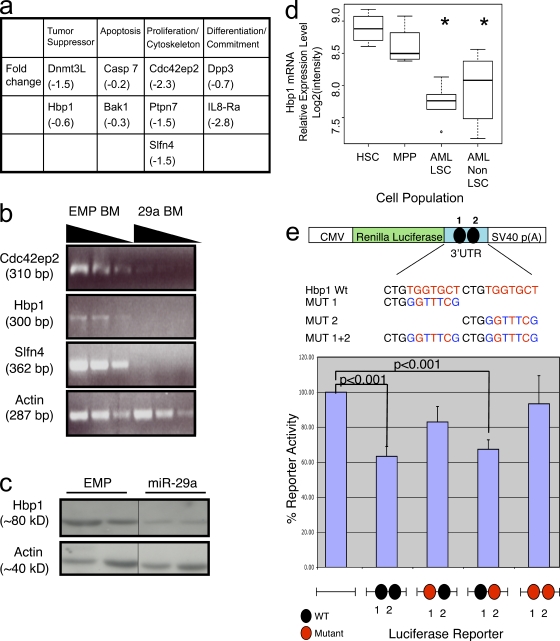
The function of microRNAs (miRNAs) in hematopoietic stem cells (HSCs), committed progenitors, and leukemia stem cells (LSCs) is poorly understood. We show that miR-29a is highly expressed in HSC and down-regulated in hematopoietic progenitors. Ectopic expression of miR-29a in mouse HSC/progenitors results in acquisition of self-renewal capacity by myeloid progenitors, biased myeloid differentiation, and the development of a myeloproliferative disorder that progresses to acute myeloid leukemia (AML). miR-29a promotes progenitor proliferation by expediting G1 to S/G2 cell cycle transitions. miR-29a is overexpressed in human AML and, like human LSC, miR-29a-expressing myeloid progenitors serially transplant AML. Our data indicate that miR-29a regulates early hematopoiesis and suggest that miR-29a initiates AML by converting myeloid progenitors into self-renewing LSC.
Figures
Similar articles
-
MicroRNA-29a and microRNA-142-3p are regulators of myeloid differentiation and acute myeloid leukemia.Blood. 2012 May 24;119(21):4992-5004. doi: 10.1182/blood-2011-10-385716. Epub 2012 Apr 4. Blood. 2012. PMID: 22493297
-
miR-29a maintains mouse hematopoietic stem cell self-renewal by regulating Dnmt3a.Blood. 2015 Apr 2;125(14):2206-16. doi: 10.1182/blood-2014-06-585273. Epub 2015 Jan 29. Blood. 2015. PMID: 25634742 Free PMC article.
-
MicroRNA-150 expression induces myeloid differentiation of human acute leukemia cells and normal hematopoietic progenitors.PLoS One. 2013 Sep 24;8(9):e75815. doi: 10.1371/journal.pone.0075815. eCollection 2013. PLoS One. 2013. PMID: 24086639 Free PMC article.
-
MicroRNAs expressed in hematopoietic stem/progenitor cells are deregulated in acute myeloid leukemias.Leuk Lymphoma. 2015 May;56(5):1466-74. doi: 10.3109/10428194.2014.955019. Epub 2015 Jan 21. Leuk Lymphoma. 2015. PMID: 25242094 Review.
-
Shp2 function in hematopoietic stem cell biology and leukemogenesis.Curr Opin Hematol. 2012 Jul;19(4):273-9. doi: 10.1097/MOH.0b013e328353c6bf. Curr Opin Hematol. 2012. PMID: 22504523 Free PMC article. Review.
Cited by
-
Micro-ribonucleic acid 29b inhibits cell proliferation and invasion and enhances cell apoptosis and chemotherapy effects of cisplatin via targeting of DNMT3b and AKT3 in prostate cancer.Onco Targets Ther. 2015 Mar 4;8:557-65. doi: 10.2147/OTT.S76484. eCollection 2015. Onco Targets Ther. 2015. PMID: 25784815 Free PMC article.
-
Reduced miR-29a-3p expression is linked to the cell proliferation and cell migration in gastric cancer.World J Surg Oncol. 2015 Mar 12;13:101. doi: 10.1186/s12957-015-0513-x. World J Surg Oncol. 2015. PMID: 25889078 Free PMC article.
-
miRNAs in acute myeloid leukemia.Oncotarget. 2017 Jan 10;8(2):3666-3682. doi: 10.18632/oncotarget.12343. Oncotarget. 2017. PMID: 27705921 Free PMC article. Review.
-
miR-29b-3p promotes progression of MDA-MB-231 triple-negative breast cancer cells through downregulating TRAF3.Biol Res. 2019 Jul 26;52(1):38. doi: 10.1186/s40659-019-0245-4. Biol Res. 2019. PMID: 31349873 Free PMC article.
-
CD97 is a critical regulator of acute myeloid leukemia stem cell function.J Exp Med. 2019 Oct 7;216(10):2362-2377. doi: 10.1084/jem.20190598. Epub 2019 Aug 1. J Exp Med. 2019. PMID: 31371381 Free PMC article.
References
-
- Abrahamsson A.E., Geron I., Gotlib J., Dao K.H., Barroga C.F., Newton I.G., Giles F.J., Durocher J., Creusot R.S., Karimi M., et al. 2009. Glycogen synthase kinase 3beta missplicing contributes to leukemia stem cell generation. Proc. Natl. Acad. Sci. USA. 106:3925–3929 10.1073/pnas.0900189106 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
