

The evolution of cancer modeling: the shadow of stem cells
- PMID: 20212083
- PMCID: PMC2869490
- DOI: 10.1242/dmm.002774
The evolution of cancer modeling: the shadow of stem cells
Abstract
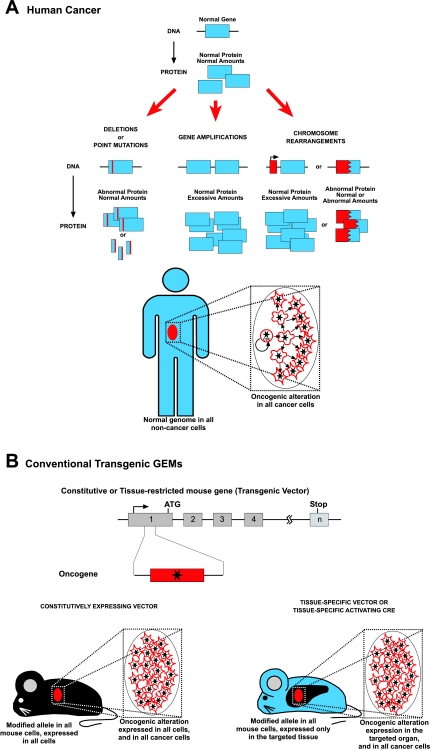
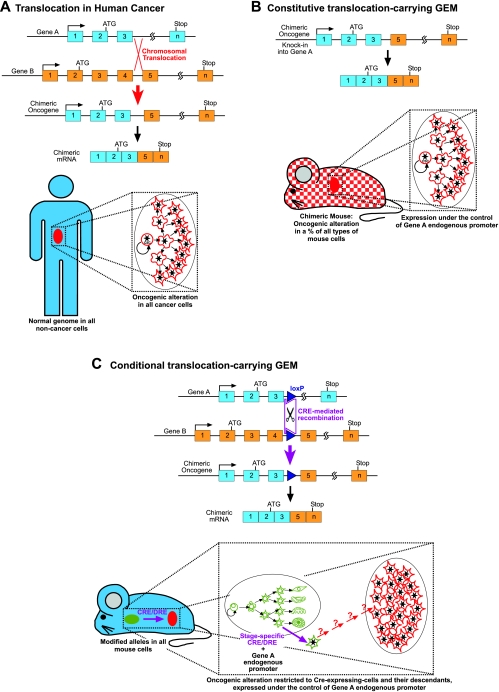
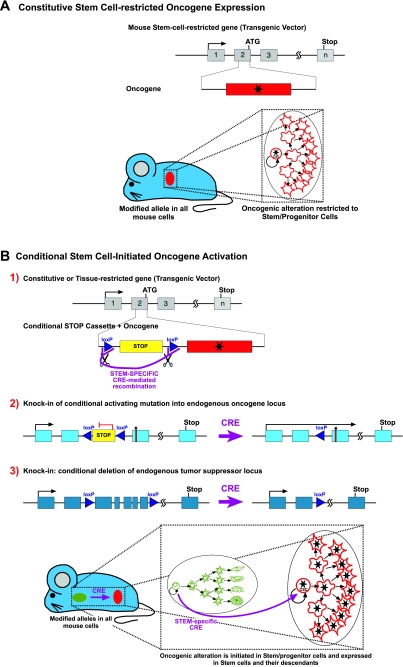
Cancer is a complex and highly dynamic process. Genetically engineered mouse models (GEMs) that develop cancer are essential systems for dissecting the processes that lead to human cancer. These animal models provide a means to determine the causes of malignancy and to develop new treatments, thus representing a resource of immense potential for medical oncology. The sophistication of modeling cancer in mice has increased to the extent that now we can induce, study and manipulate the cancer disease process in a manner that is impossible to perform in human patients. However, all GEMs described so far have diverse shortcomings in mimicking the hierarchical structure of human cancer tissues. In recent years, a more detailed picture of the cellular and molecular mechanisms determining the formation of cancer has emerged. This Commentary addresses new experimental approaches toward a better understanding of carcinogenesis and discusses the impact of new animal models.
Figures
References
-
- Adams JM, Harris AW, Pinkert CA, Corcoran LM, Alexander WS, Cory S, Palmiter RD, Brinster RL. (1985). The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature 318, 533–538 - PubMed
-
- Ailles LE, Weissman IL. (2007). Cancer stem cells in solid tumors. Curr Opin Biotechnol. 18, 460–466 - PubMed
-
- Anastassiadis K, Fu J, Patsch C, Hu S, Weidlich S, Duerschke K, Buchholz F, Edenhofer F, Stewart AF. (2009). Dre recombinase, like Cre, is a highly efficient site-specific recombinase in E. coli, mammalian cells and mice. Dis Model Mech. 2, 508–515 - PubMed
-
- Barker N, Ridgway RA, van Es JH, van de Wetering M, Begthel H, van den Born M, Danenberg E, Clarke AR, Sansom OJ, Clevers H. (2009). Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 457, 608–611 - PubMed
