

Review
doi: 10.1021/cr900228f.
Mechanisms for DNA charge transport
Affiliations
- PMID: 20214403
- PMCID: PMC2879062
- DOI: 10.1021/cr900228f
Item in Clipboard
Review
Mechanisms for DNA charge transport
Chem Rev.
.
No abstract available
Figures
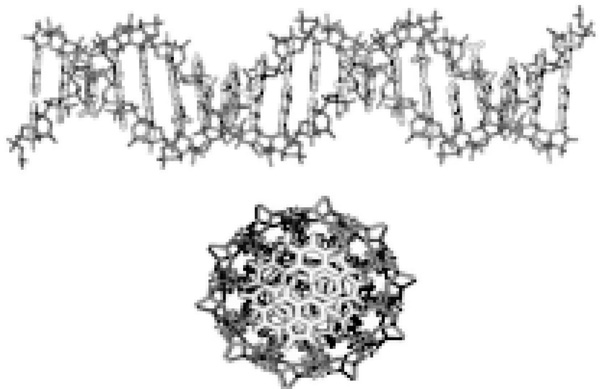
Transverse and longitudinal perspectives of DNA. The sugar phosphate backbone envelops the hydrophobic base pairs. The planar base pairs form a one-dimensional π-stack down the center of the DNA, insulated by the backbone.
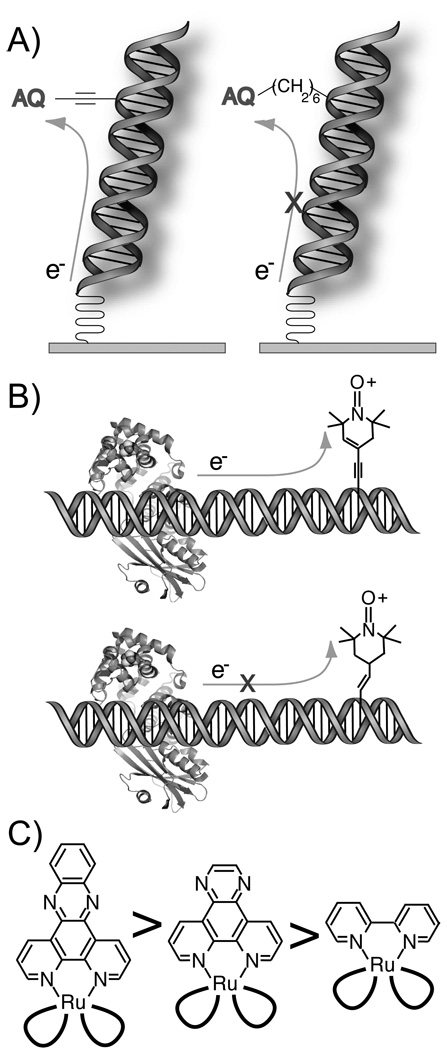
DNA-mediated CT requires electronic coupling to the base pair stack. (A) Electrochemical reduction of an electronically well-coupled anthraquinone (AQ) is facile, while that of a poorly coupled AQ is suppressed. (B) MutY competently reduces an oxidized nitroxide spin label that is well coupled to thymidine, but not the nitroxide conjugated through the partially unsaturated linker. (C) For a series of polypyridyl RuIII(bpy)2L ground state oxidants, the yield of oxidative damage to DNA scales with the size and planarity of the intercalating ligand.
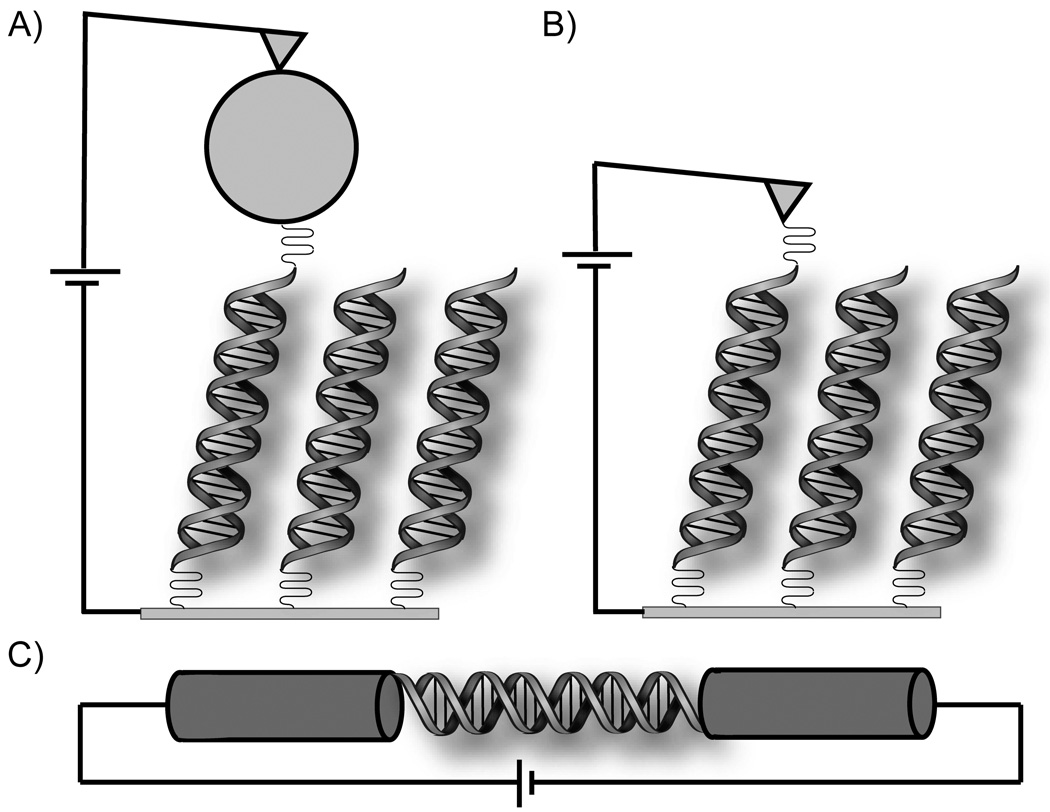
Devices for measurement of single molecule DNA conductivity through molecular contacts. In each case, currents between 10 and 100 nA are obtained for modest source-drain and gating voltages. A) A gold nanoparticle allows strong coupling between the EC-AFM tip and an individual 26mer DNA molecule on a gold electrode. B) The gold STM tip is slowly brought in contact with thiol-modified DNA (8mer), allowing a histogram of conductance over many different orientations. C) A single 15mer DNA is covalently attached across a gap between single-walled carbon nanotubes.
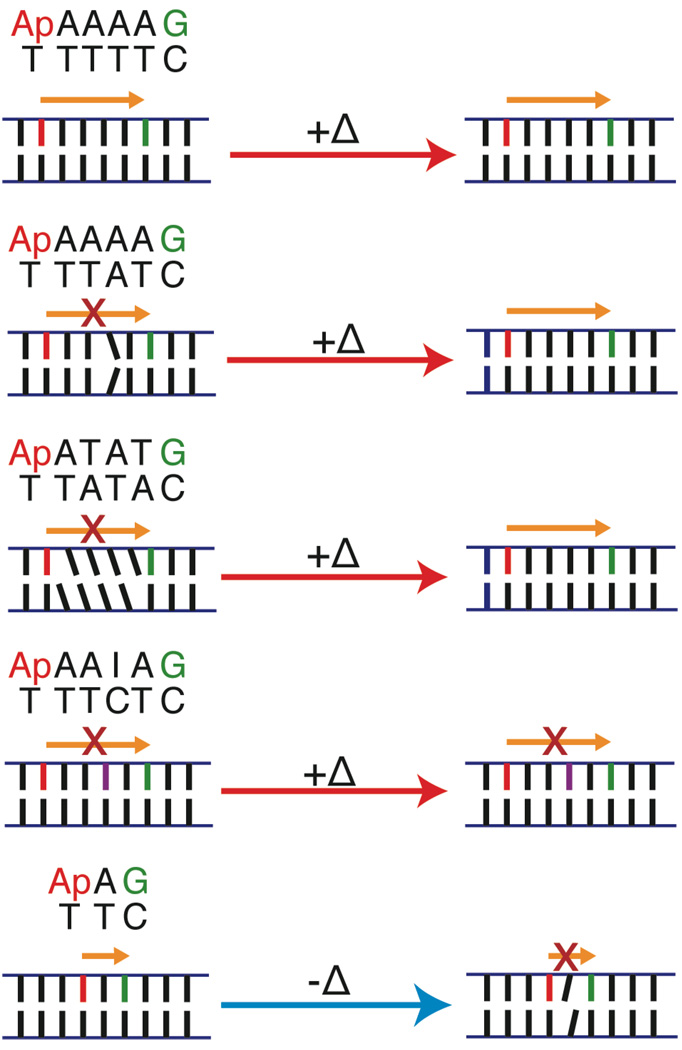
The sequence and temperature dependence of single-step oxidation of guanine by photoexcited 2-aminopurine (Ap) in DNA.,, CT yield in well-matched Ap(A)4G increases with temperature (+Δ), up to duplex melting. Two perturbations that disturb CT due to poor stacking dynamics, an A–A mismatch and the sequence ATAT, attenuate CT at room temperature but are comparable to the A4 sequence at higher temperature, while CT through a perturbation that disturbs CT due to an electronic barrier, AAIA, is only partially recovered at high temperature. This argues that the CT activation is related to the flexibility of the bridge. At low temperature (77K), an intervening adenine eliminates CT from Ap to G, implying that the equilibrium conformation is not the CT active conformation.
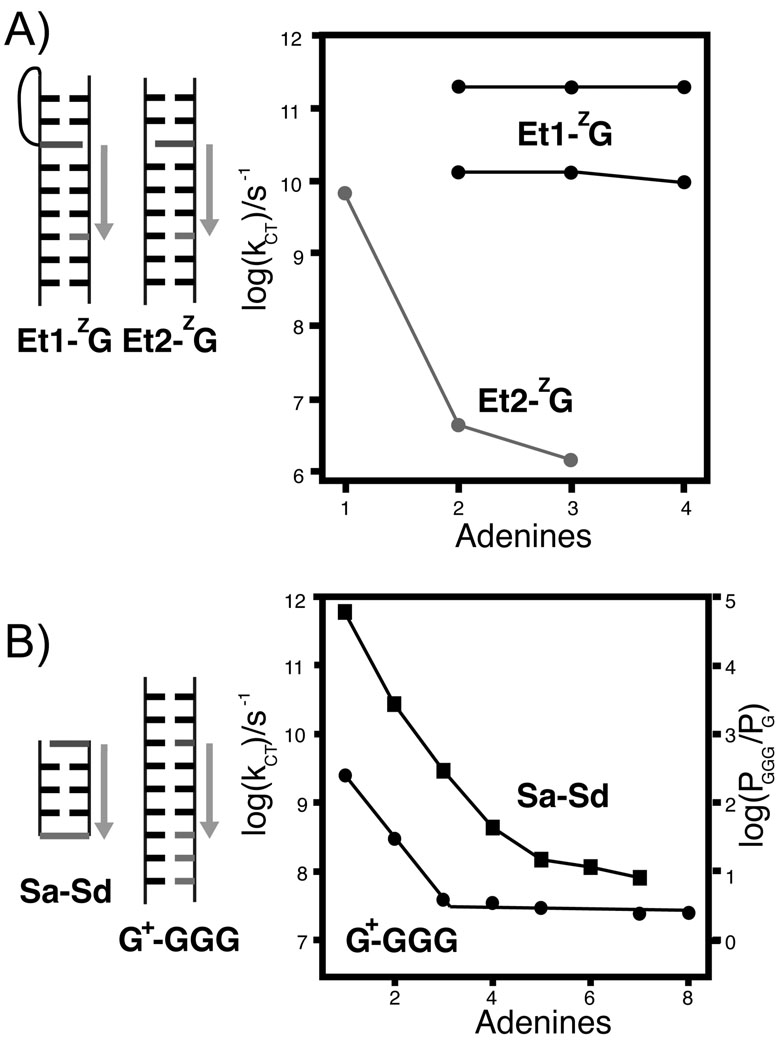
(A) The rate of coherent deazaguanine (ZG) oxidation by ethidium bromide is the same over short distances for both the flexible linkage (Et1, black, both CT rates shown) and the rigid linkage (Et2, gray). For two intervening nucleotides, a sharp drop in rate is observed for the rigid Et+, but the rate is unaffected for the flexible Et+. (B) This steep drop in rate over short distances is consistent with that observed for oxidation of guanine by a photoinduced sugar radical (circles), and CT between hairpin capping stilbenes (squares, photooxidation of Sd by Sa), and has been attributed to a crossover between coherent superexchange and incoherent hopping. In the latter case, comparison of injection and hole arrival rates supports superexchange for one or two intervening base pairs, and hopping for three or more base pairs between the stilbenes.
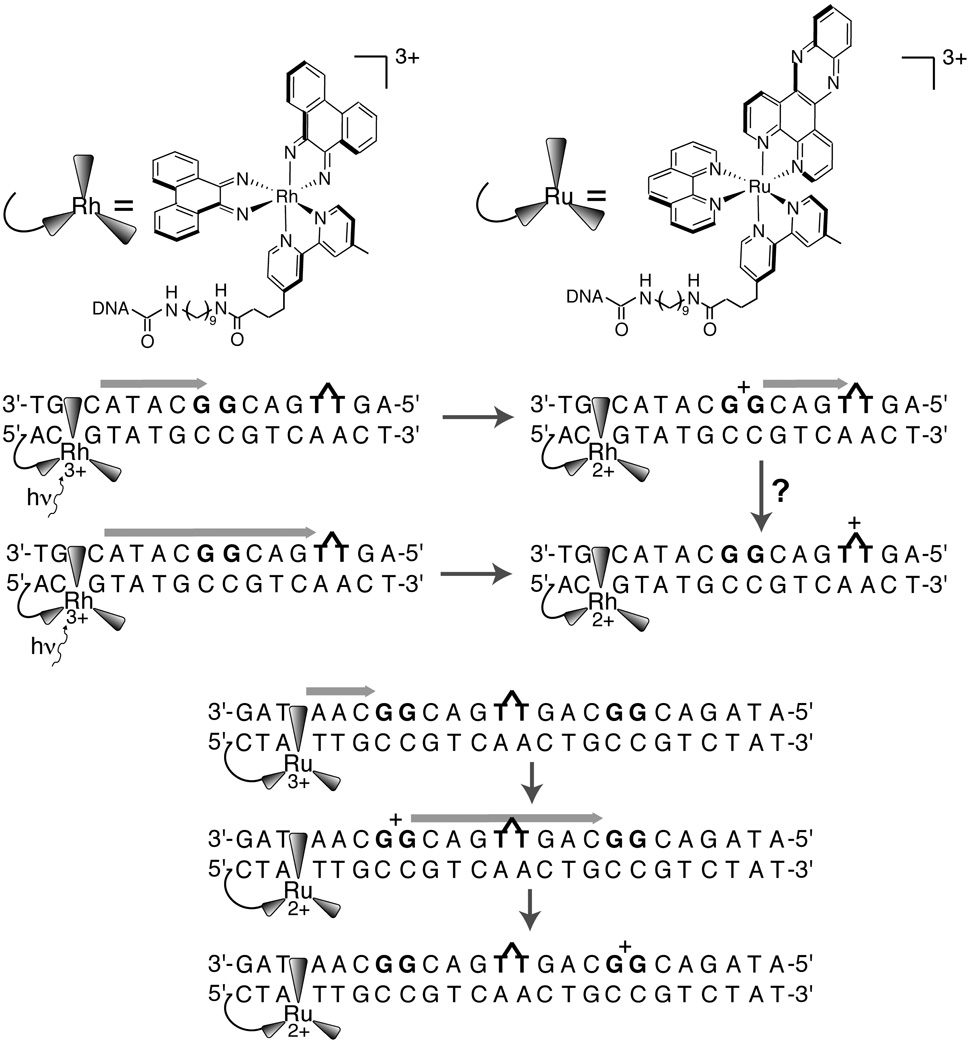
Oxidation and repair of thymine dimer (~1.8 eV) by tethered photoexcited [Rh(phi)2(bpy’)]3+ (2.0 V) is unaffected by the intervening double guanine site (1.2 eV). Oxidation of double guanine sites by [Ru(phen)(bpy’)(dppz)]3+ (1.5 eV) is unaffected by the presence of thymine dimer, which this oxidant lacks sufficient driving force to repair. The latter result implies that guanine radical is not competent to repair thymine dimers, in accordance with the known potentials. Hence either the guanine radical oxidized by [Rh(phi)2(bpy’)]3+ does not relax prior to migration to the thymine dimer, or the guanine radical is not an intermediate in DNA-mediated oxidation of thymine dimer by [Rh(phi)2(bpy’)]3+.
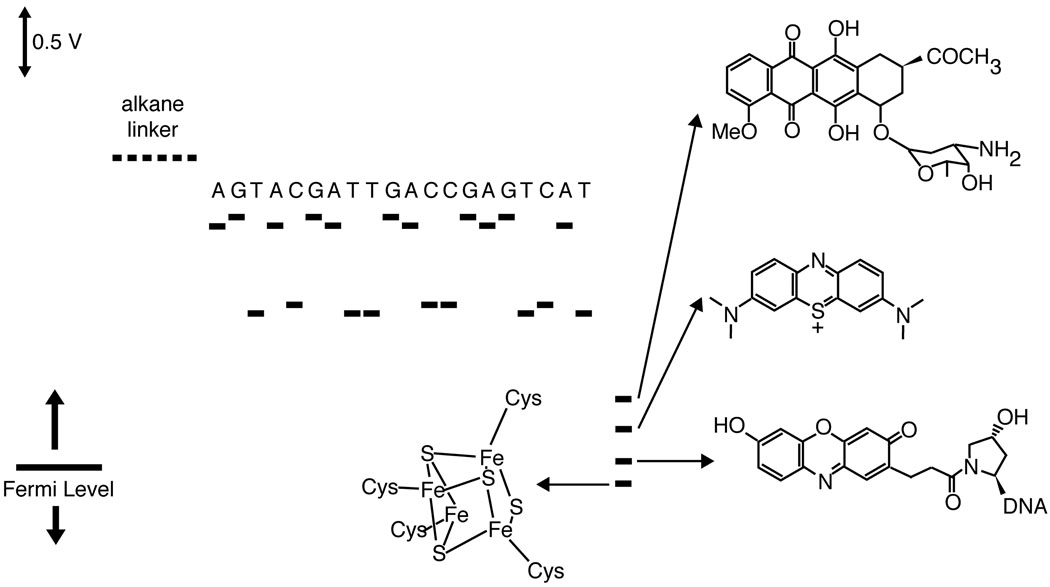
Scale diagram describing the relevant potentials for DNA-mediated CT through DNA self-assembled monolayers on gold. The potentials of the individual nucleotides are not accessible within the window of electrochemistry of DNA monolayers on Au. Nevertheless, facile DNA-mediated electrochemistry is observed for redox probes over DNA bridges. For all probes and sequences of well-matched duplexes, the tunneling through the alkane linker is rate limiting (~30 s−1). Shown, in order from top, are daunomycin, methylene blue, Redmond Red, and a [4Fe-4S] cluster similar to those in the redox-active repair proteins EndoIII and MutY.
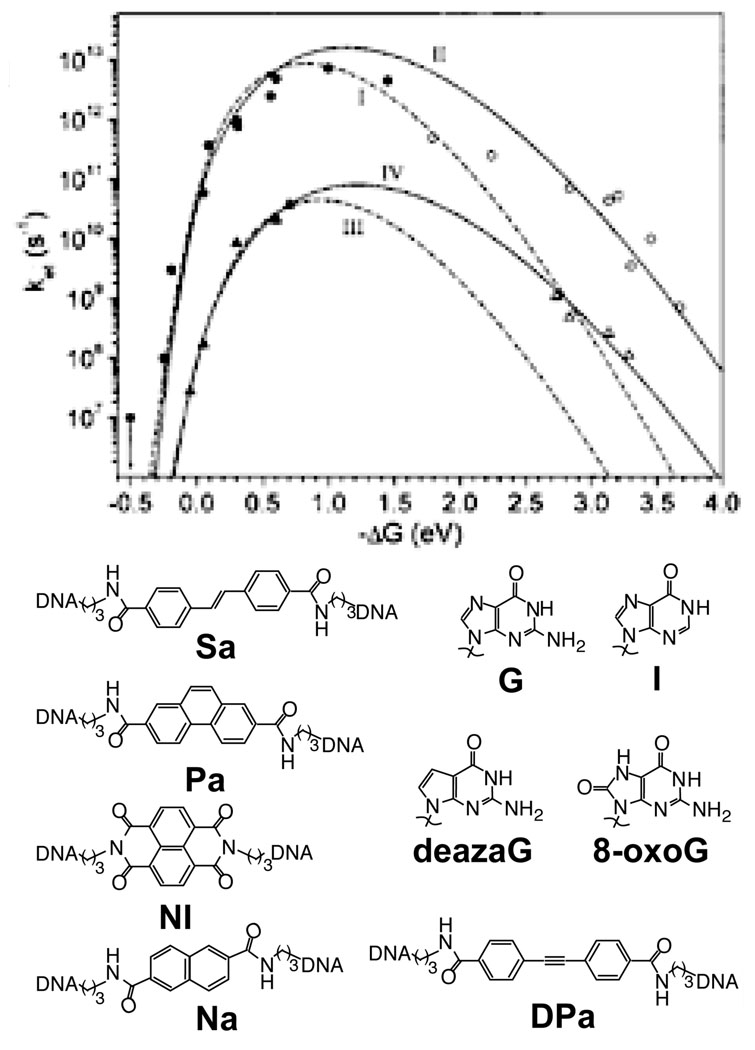
The driving force dependence for CT in photooxidant-bridged DNA hairpins is determined from time-resolved transient absorption studies of a series of five stilbene-derived photooxidants and four hole acceptor bases, following both charge separation (filled) and charge recombination (empty). Case I,II (circles): donor and acceptor are in contact. Case III,IV (triangles): donor and acceptor are separated by two TA base pairs. Case I,III are fit only to charge separation rates (dotted), while Case II,IV are fit to both charge separation and charge recombination rates (solid). Similar reorganization energies, about 1 eV for the nuclear reorganization energy and ~0.2 V for the solvent reorganization energy, are found for both Case II and Case IV. Reprinted with permission from Reference . Copyright American Chemical Society.
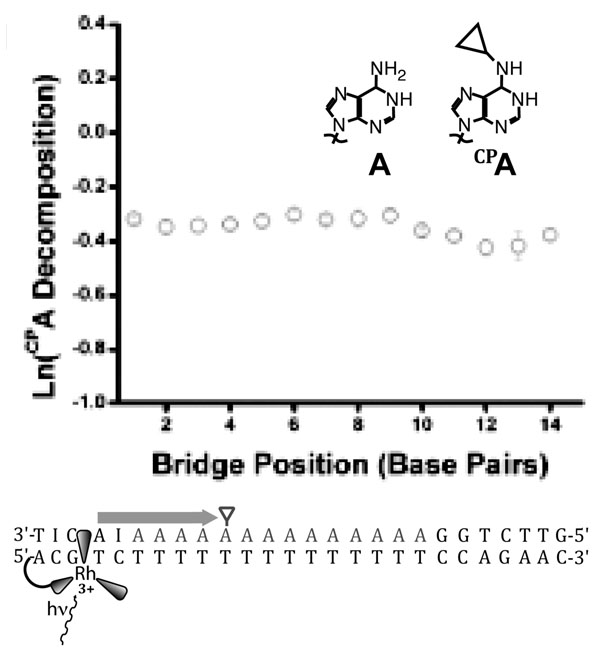
CT from photoexcited [Rh(phi)2(bpy’)]3+ to N-cyclopropyladenosine (CPA) across an adenine tract is distance-independent over 14 adenines. The rate of CT across the adenine tract, then, must be much faster than BET from the first adenine to the reduced rhodium. The driving force for recombination is only about 1.7 V, implying that BET should not be in the inverted region, consistent with evidence that BET from adenine to this rhodium complex is facile. The lack of distance-dependence, in a system with a rapid competing process in BET and a charge trap that samples pre-equilibrium CT dynamics, implies extensive delocalization across the bridge.
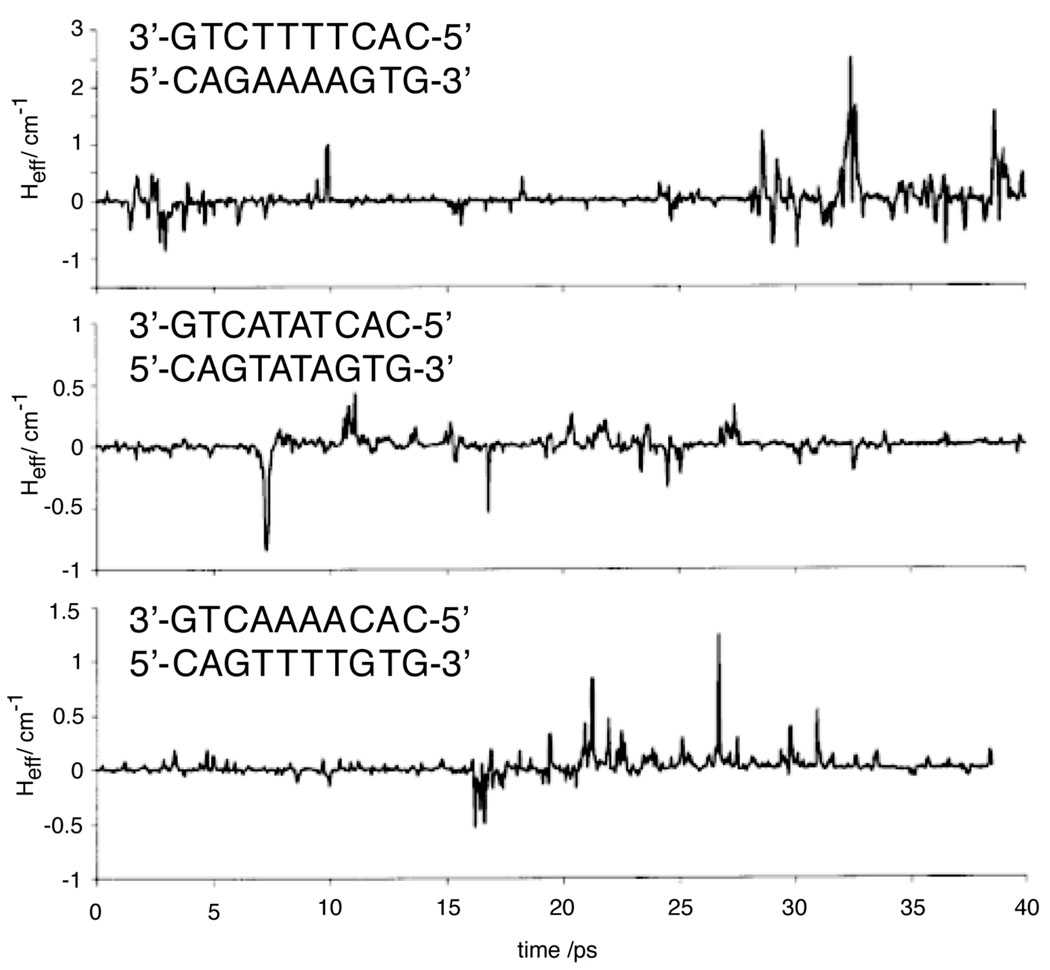
The time-dependent couplings between guanines separated by three different four-base sequence contexts, based on conformations generated through molecular dynamics. It is clear that the average value of coupling can be several orders of magnitude lower than the maximum coupling. For the poorly stacked, flexible ATAT sequence, strong coupling between the guanines is not achieved over the time-scale of the simulation. Reprinted with permission from Reference . Copyright American Chemical Society.
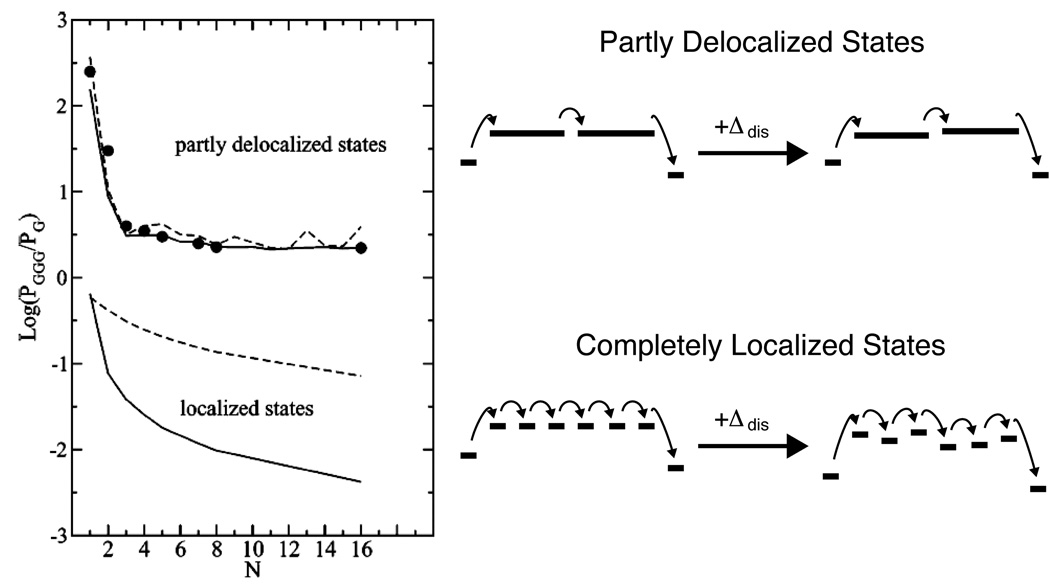
The variable range hopping model predicts a shallow distance dependence for the rate of CT between G and GGG across an adenine tract of the opposing strand. Delocalized states, even in the absence of disorder (dashed lines), yield larger and shallower CT rates due to the smaller reorganization energy, and a shorter effective bridge length. In the presence of static disorder (solid lines), localized hopping is substantially attenuated due to the rugged energy landscape. Delocalized hopping, however, is relatively unaffected by static disorder, as the coupling is strong enough to allow tunneling through local barriers. Reprinted from Reference with permission. Copyright American Chemical Society.
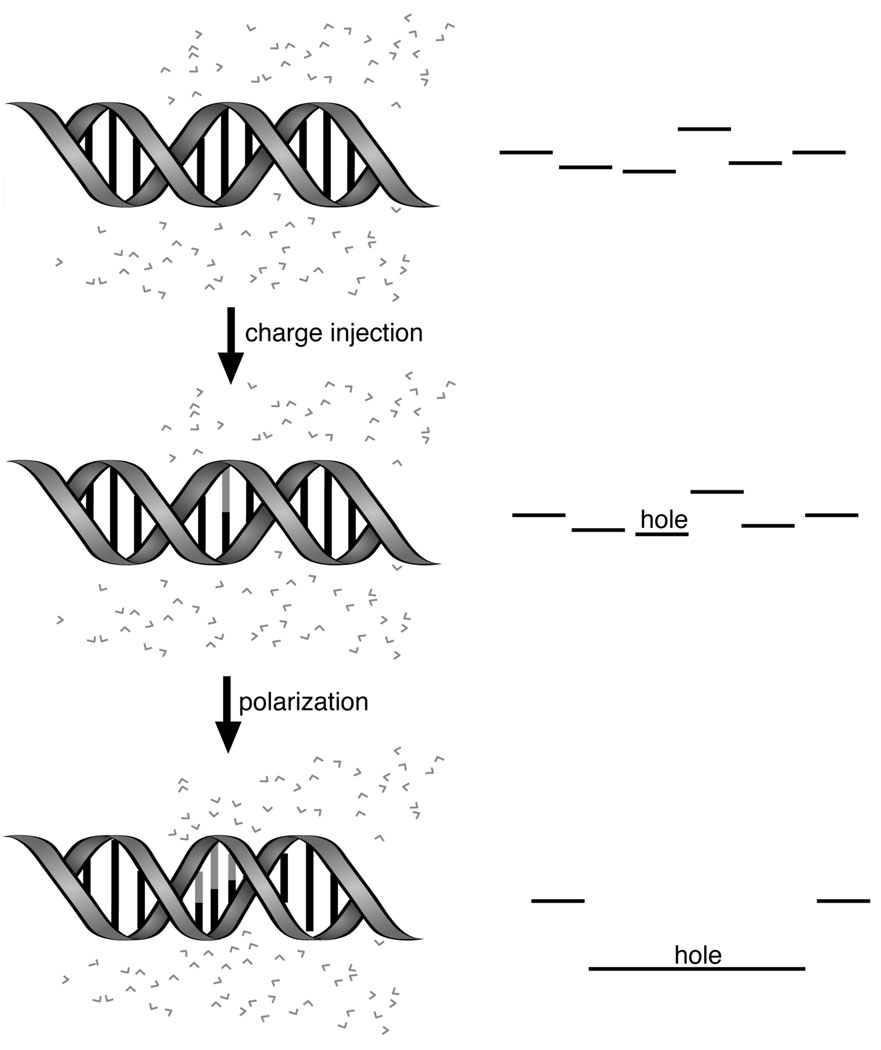
Formation of a large polaron. Upon charge injection, a hole is initially localized on a single base (red). Reorientation of the environment, including neighboring bases and the hydration layer, lowers the energy of the hole. Delocalization occurs to the extent that the coupling between the bases balances the unfavorable decrease in the reorganization energy.
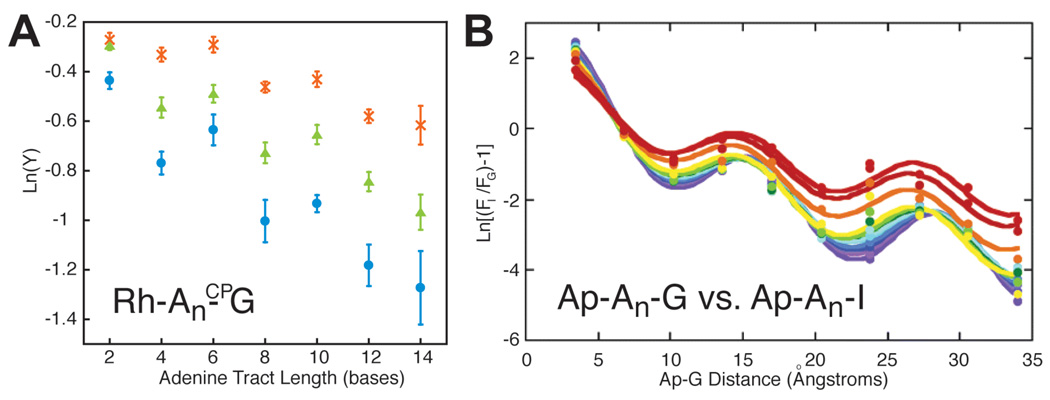
Equivalent periodicities with the same period and temperature dependence are observed for (B) the single-step oxidation of guanine by Ap* and (A) the total oxidation of CPG by photoexcited [Rh(phi)2(bpy’)]3+. Temperature increases from purple to red. Errors are given in (A) as 90% SEM., Reprinted with permission from References and . Copyright American Chemical Society.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
