

Attenuation of Mycobacterium tuberculosis functionally disrupted in a fatty acyl-coenzyme A synthetase gene fadD5
- PMID: 20214478
- PMCID: PMC3225055
- DOI: 10.1086/651452
Attenuation of Mycobacterium tuberculosis functionally disrupted in a fatty acyl-coenzyme A synthetase gene fadD5
Abstract
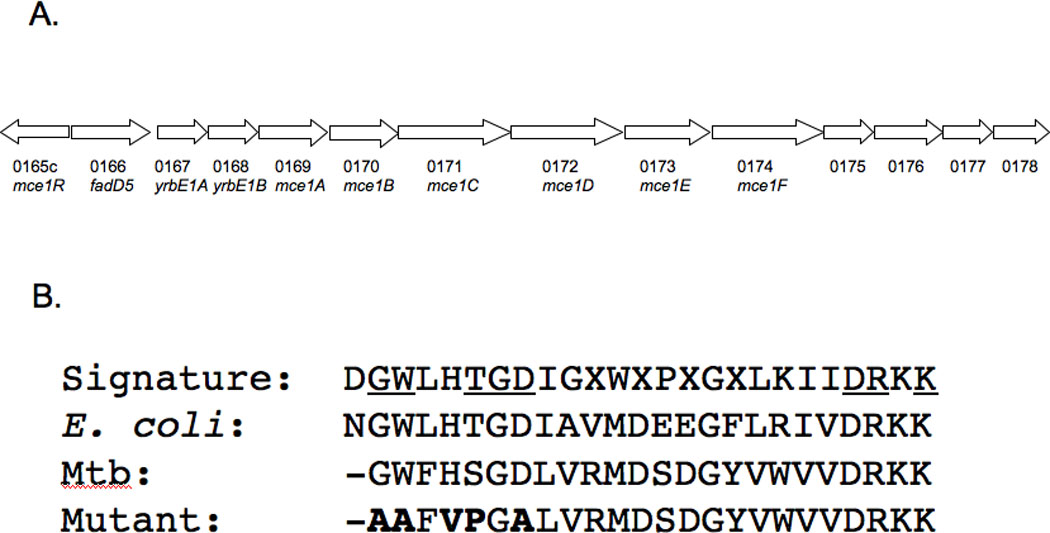
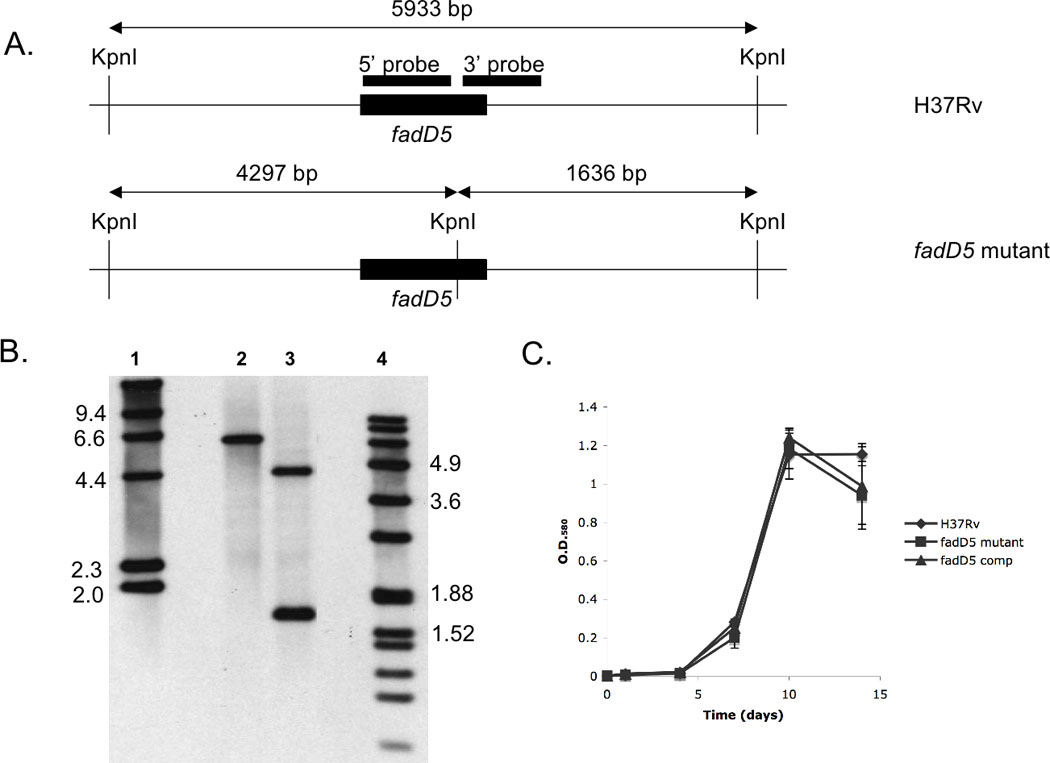
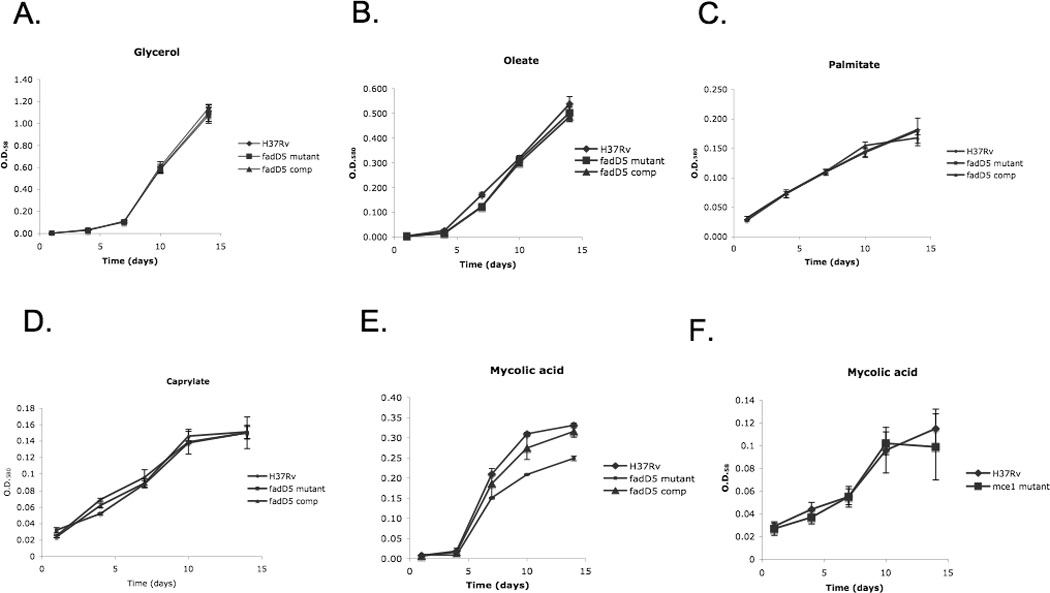
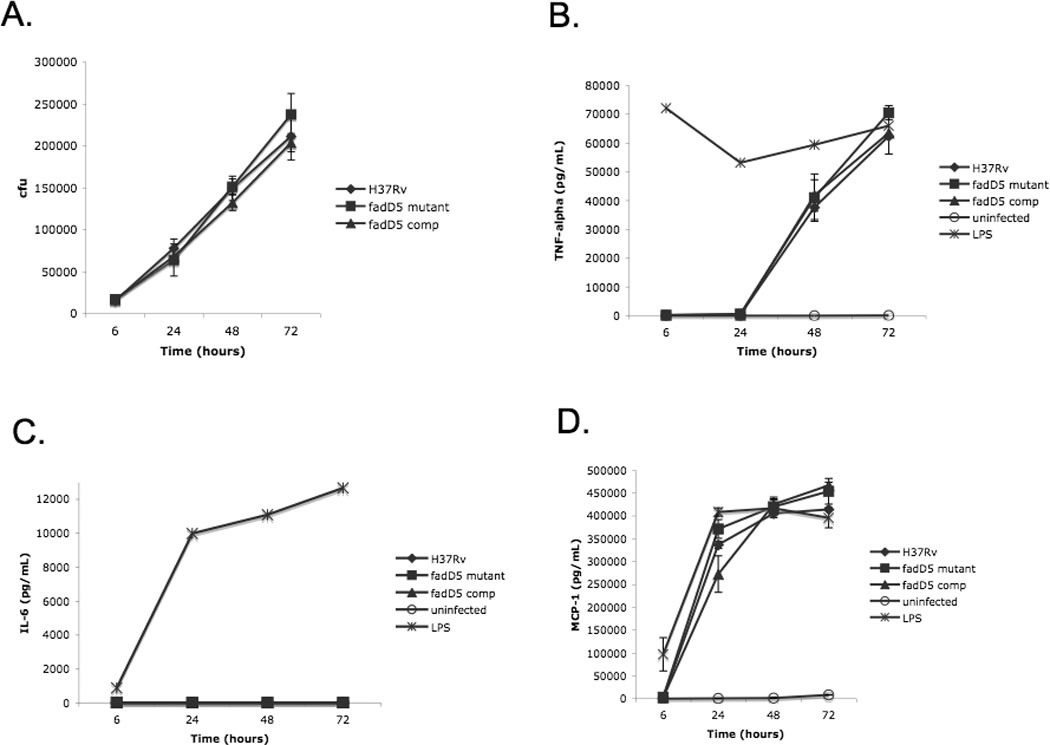
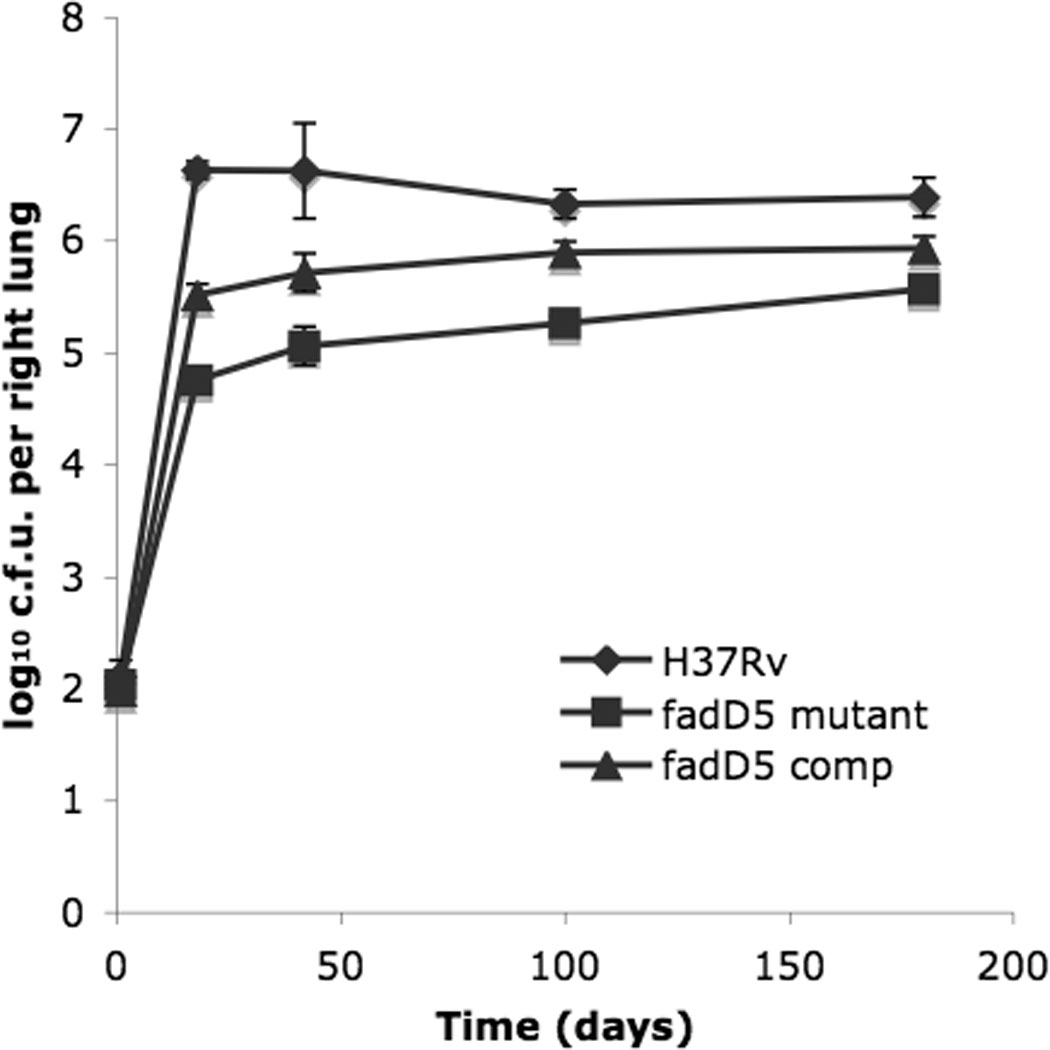
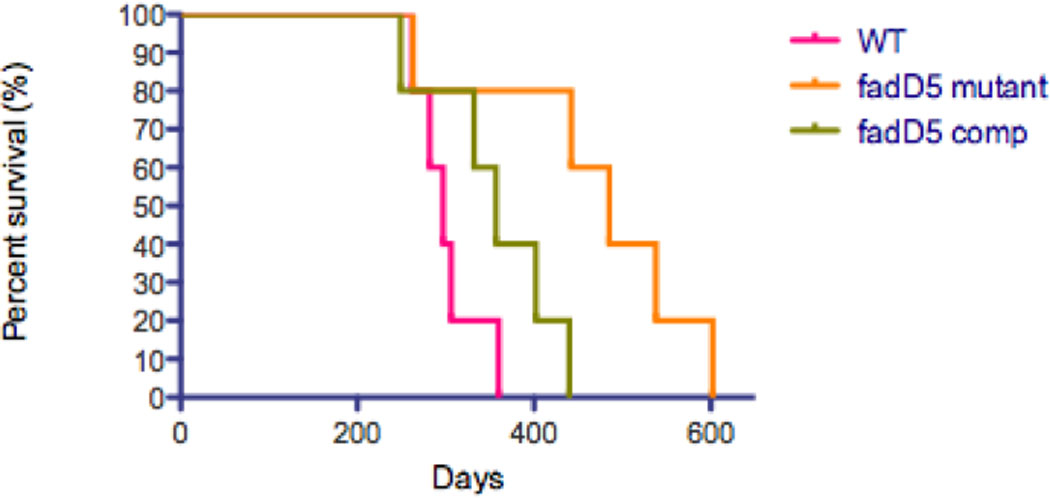
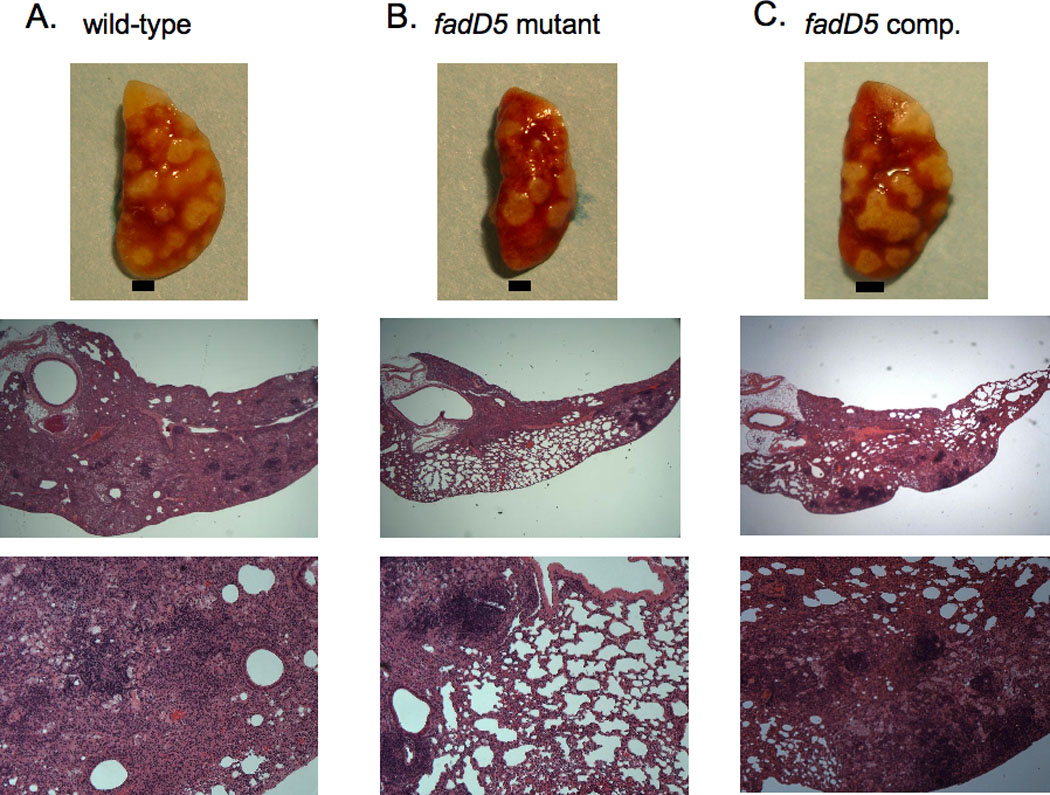
One key adaptation that Mycobacterium tuberculosis established to survive long term in vivo is a reliance on lipids as an energy source. M. tuberculosis H37Rv has 36 fadD genes annotated as putative fatty acyl-coenzyme A (CoA) synthetase genes, which encode enzymes that activate fatty acids for metabolism. One such gene, fadD5 (Rv0166), is located within the mce1 operon, a cluster of genes associated with M. tuberculosis persistence. We disrupted the putative fatty acid-binding site of fadD5 in H37Rv M. tuberculosis. No significant differences were found in the growth of the mutant and wild-type strains in vitro in nutrient-rich broth or in activated RAW264.7 cells. However, the fadD5 mutant was diminished in growth in minimal medium containing mycolic acid but not other long-chain fatty acids. C57BL/6 mice infected with the fadD5 mutant survived significantly longer than those infected with the wild type, and the mutant never attained the plateau phase of infection in mouse lungs. Infection in the steady-state phase was maintained for up to 168 days at a level that was 1-2 logs less than that noted in the wild type. These observations raise the rather intriguing possibility that FadD5 may serve to recycle mycolic acids for the long-term survival of the tubercle bacilli.
Conflict of interest statement
The authors do not have a commercial or other association that might post a conflict of interest (e.g., pharmaceutical stock ownership, consultancy, advisory board membership, relevant patents, or research funding).
Figures
References
-
- Parrish NM, Dick JD, Bishai WR. Mechanisms of latency in Mycobacterium tuberculosis. Trends Microbiol. 1998;6:107–112. - PubMed
-
- Corbett EL, Watt CJ, Walker N, et al. The growing burden of tuberculosis: global trends and interactions with the HIV epidemic. Arch Intern Med. 2003;163:1009–1021. - PubMed
-
- Dye C, Scheele S, Dolin P, Pathania V, Raviglione MC. Consensus statement. Global burden of tuberculosis: estimated incidence, prevalence, and mortality by country. WHO Global Surveillance and Monitoring Project. JAMA. 1999;282:677–686. - PubMed
-
- Betts JC, Lukey PT, Robb LC, McAdam RA, Duncan K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol Microbiol. 2002;43:717–731. - PubMed
-
- Cole ST, Brosch R, Parkhill J, et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature. 1998;393:537–544. - PubMed
