

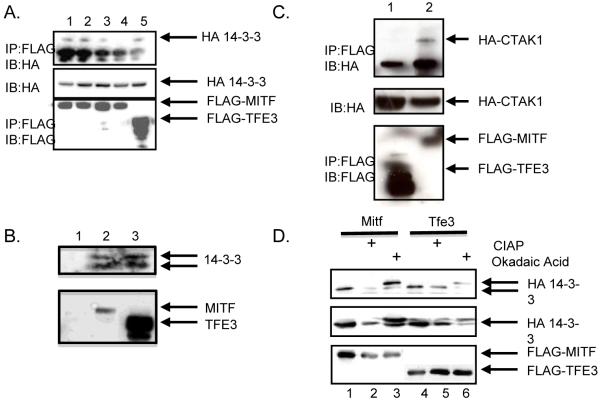
C-TAK1 interacts with microphthalmia-associated transcription factor, Mitf, but not the related family member Tfe3
- PMID: 20214879
- PMCID: PMC2858567
- DOI: 10.1016/j.bbrc.2010.03.034
C-TAK1 interacts with microphthalmia-associated transcription factor, Mitf, but not the related family member Tfe3
Abstract
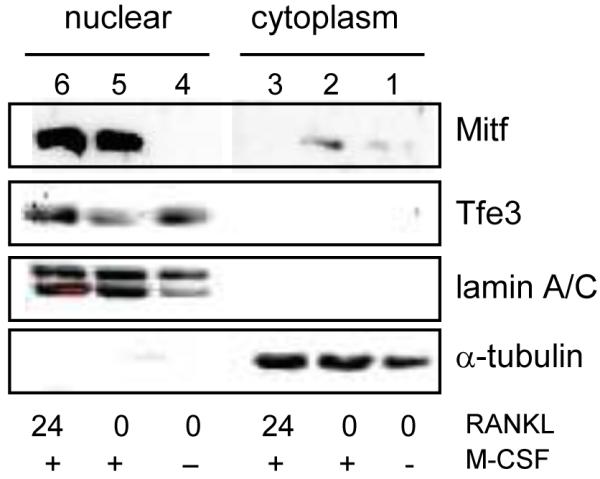
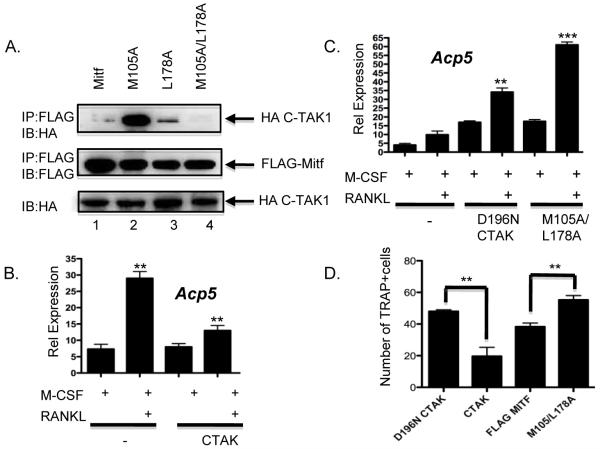
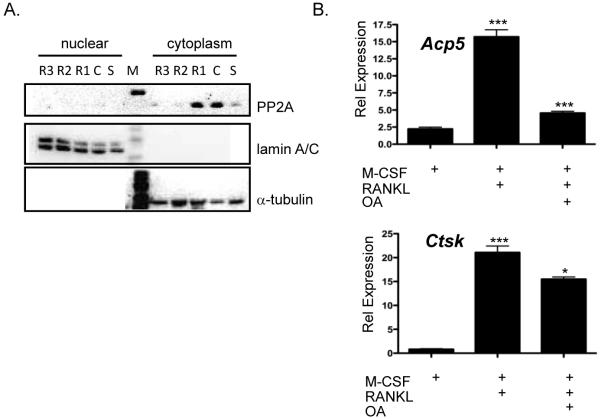
Microphthalmia-associated transcription factor, Mitf, has been shown to be necessary for regulating genes involved in osteoclast differentiation. Previously it was shown by others that Mitf translocates from the cytoplasm to the nucleus upon M-CSF/RANKL signaling in osteoclasts. Mitf's movement is regulated by its interaction with 14-3-3 and the kinase C-TAK1. Here we demonstrate that the related family member, Tfe3, does not shuttle from the cytoplasm to the nucleus and does not interact with C-TAK1. We also demonstrate that overexpression of C-TAK1 inhibits the expression of Acp5 while a kinase dead C-TAK1 or a Mitf mutant that cannot interact with C-TAK1 increased expression of Acp5. Finally, we show that the catalytic subunit of protein phosphatase 2A is up-regulated in osteoclasts with M-CSF/RANKL signaling, indicating a possible mechanism for dephosphorylating Mitf on its 14-3-3 binding site and allowing Mitf to be translocated to the nucleus of osteoclasts.
2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Hemesath TJ, Steingrimsson E, McGill G, Hansen MJ, Vaught J, Hodgkinson CA, Arnheiter H, Copeland NG, Jenkins NA, Fisher DE. microphthalmia, a critical factor in melanocyte development, defines a discrete transcription factor family. Genes Dev. 1994;8:2770–80. - PubMed
-
- Hershey CL, Fisher DE. Mitf and Tfe3: members of a b-HLH-ZIP transcription factor family essential for osteoclast development and function. Bone. 2004;34:689–96. - PubMed
-
- Steingrimsson E, Copeland NG, Jenkins NA. Melanocytes and the microphthalmia transcription factor network. Annu Rev Genet. 2004;38:365–411. - PubMed
-
- Luchin A, Purdom G, Murphy K, Clark MY, Angel N, Cassady AI, Hume DA, Ostrowski MC. The microphthalmia transcription factor regulates expression of the tartrate-resistant acid phosphatase gene during terminal differentiation of osteoclasts. J Bone Miner Res. 2000;15:451–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
