

PHOX2A regulation of oculomotor complex nucleogenesis
- PMID: 20215354
- PMCID: PMC2835333
- DOI: 10.1242/dev.041251
PHOX2A regulation of oculomotor complex nucleogenesis
Abstract
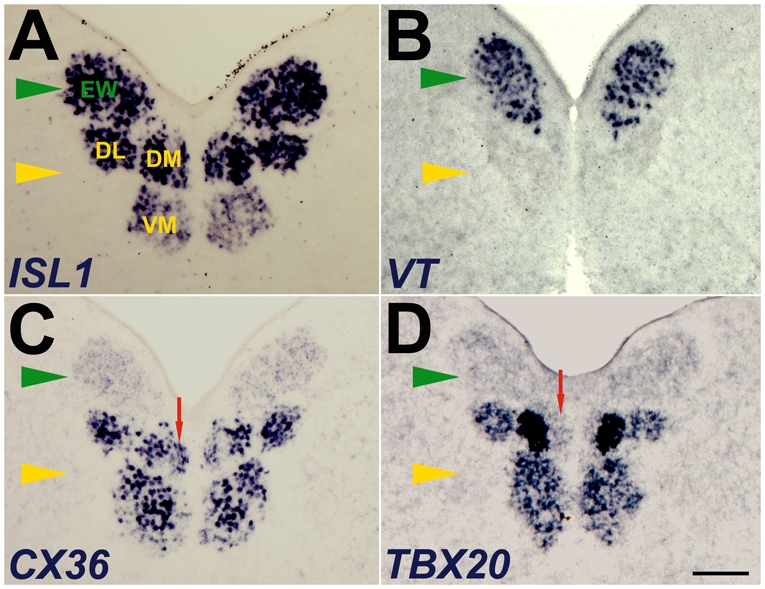
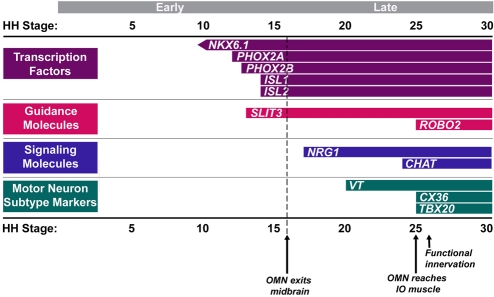
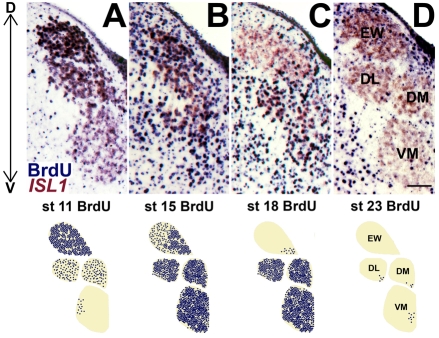
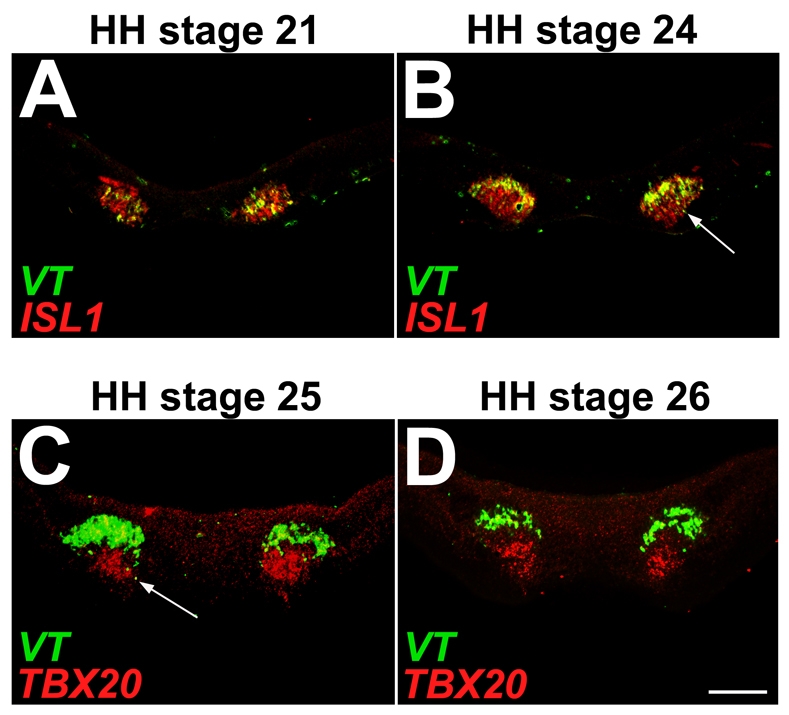
Brain nuclei are spatially organized collections of neurons that share functional properties. Despite being central to vertebrate brain circuitry, little is known about how nuclei are generated during development. We have chosen the chick midbrain oculomotor complex (OMC) as a model with which to study the developmental mechanisms of nucleogenesis. The chick OMC comprises two distinct cell groups: a dorsal Edinger-Westphal nucleus of visceral oculomotor neurons and a ventral nucleus of somatic oculomotor neurons. Genetic studies in mice and humans have established that the homeobox transcription factor gene PHOX2A is required for midbrain motoneuron development. We probed, in forced expression experiments, the capacity of PHOX2A to generate a spatially organized midbrain OMC. We found that exogenous Phox2a delivery to embryonic chick midbrain can drive a complete OMC molecular program, including the production of visceral and somatic motoneurons. Phox2a overexpression was also able to generate ectopic motor nerves. The exit points of such auxiliary nerves were invested with ectopic boundary cap cells and, in four examples, the ectopic nerves were seen to innervate extraocular muscle directly. Finally, Phox2a delivery was able to direct ectopic visceral and somatic motoneurons to their correct native spatial positions, with visceral motoneurons settling close to the ventricular surface and somatic motoneurons migrating deeper into the midbrain. These findings establish that in midbrain, a single transcription factor can both specify motoneuron cell fates and orchestrate the construction of a spatially organized motoneuron nuclear complex.
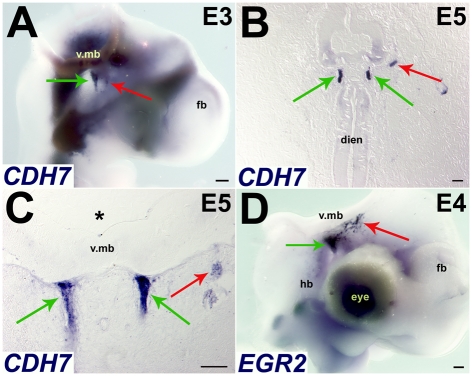
Figures
Similar articles
-
Nkx6-1 controls the identity and fate of red nucleus and oculomotor neurons in the mouse midbrain.Development. 2009 Aug;136(15):2545-55. doi: 10.1242/dev.031781. Development. 2009. PMID: 19592574 Free PMC article.
-
Complementary roles for Nkx6 and Nkx2 class proteins in the establishment of motoneuron identity in the hindbrain.Development. 2003 Sep;130(17):4149-59. doi: 10.1242/dev.00641. Development. 2003. PMID: 12874134
-
A role for midbrain arcs in nucleogenesis.Development. 2002 Dec;129(24):5779-88. doi: 10.1242/dev.00179. Development. 2002. PMID: 12421716
-
The genetic basis of complex strabismus.Pediatr Res. 2006 Mar;59(3):343-8. doi: 10.1203/01.pdr.0000200797.91630.08. Pediatr Res. 2006. PMID: 16492969 Review.
-
Turning heads: development of vertebrate branchiomotor neurons.Dev Dyn. 2004 Jan;229(1):143-61. doi: 10.1002/dvdy.10444. Dev Dyn. 2004. PMID: 14699587 Free PMC article. Review.
Cited by
-
Extraocular motoneuron pools develop along a dorsoventral axis in zebrafish, Danio rerio.J Comp Neurol. 2017 Jan 1;525(1):65-78. doi: 10.1002/cne.24042. Epub 2016 Jun 1. J Comp Neurol. 2017. PMID: 27197595 Free PMC article.
-
Motor neurons are dispensable for the assembly of a sensorimotor circuit for gaze stabilization.bioRxiv [Preprint]. 2024 Jan 27:2024.01.25.577261. doi: 10.1101/2024.01.25.577261. bioRxiv. 2024. Update in: Elife. 2024 Nov 20;13:RP96893. doi: 10.7554/eLife.96893. PMID: 38328255 Free PMC article. Updated. Preprint.
-
Intrinsic properties guide proximal abducens and oculomotor nerve outgrowth in avian embryos.Dev Neurobiol. 2012 Feb;72(2):167-85. doi: 10.1002/dneu.20948. Dev Neurobiol. 2012. PMID: 21739615 Free PMC article.
-
Multi-omic analysis of selectively vulnerable motor neuron subtypes implicates altered lipid metabolism in ALS.Nat Neurosci. 2021 Dec;24(12):1673-1685. doi: 10.1038/s41593-021-00944-z. Epub 2021 Nov 15. Nat Neurosci. 2021. PMID: 34782793 Free PMC article.
-
Ocular Motor Nerve Development in the Presence and Absence of Extraocular Muscle.Invest Ophthalmol Vis Sci. 2017 Apr 1;58(4):2388-2396. doi: 10.1167/iovs.16-21268. Invest Ophthalmol Vis Sci. 2017. PMID: 28437527 Free PMC article.
References
-
- Agarwala S., Ragsdale C. W. (2002). A role for midbrain arcs in nucleogenesis. Development 129, 5779-5788 - PubMed
-
- Agarwala S., Ragsdale C. W. (2009). Midbrain patterning. In Encyclopedia of Neuroscience, Vol. 5 (ed. Squire L. R.), pp. 879-888 Amsterdam: Elsevier;
-
- Agarwala S., Sanders T. A., Ragsdale C. W. (2001). Sonic Hedgehog control of size and shape in midbrain pattern formation. Science 291, 2147-2150 - PubMed
-
- Bayly R. D., Ngo M., Aglyamova G. V., Agarwala S. (2007). Regulation of ventral midbrain patterning by Hedgehog signaling. Development 134, 2115-2124 - PubMed
-
- Bosley T. M., Oystreck D. T., Robertson R. L., al Awad A., Abu-Amero K., Engle E. C. (2006). Neurological features of congenital fibrosis of the extraocular muscles type 2 with mutations in PHOX2A. Brain 129, 2363-2374 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
