

Telomeric protein TRF2 protects Holliday junctions with telomeric arms from displacement by the Werner syndrome helicase
- PMID: 20215438
- PMCID: PMC2896529
- DOI: 10.1093/nar/gkq144
Telomeric protein TRF2 protects Holliday junctions with telomeric arms from displacement by the Werner syndrome helicase
Abstract
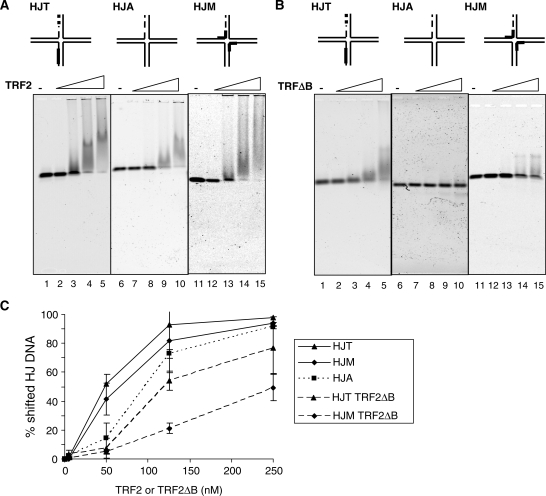
WRN protein loss causes Werner syndrome (WS), which is characterized by premature aging as well as genomic and telomeric instability. WRN prevents telomere loss, but the telomeric protein complex must regulate WRN activities to prevent aberrant telomere processing. Telomere-binding TRF2 protein inhibits telomere t-loop deletion by blocking Holliday junction (HJ) resolvase cleavage activity, but whether TRF2 also modulates HJ displacement at t-loops is unknown. In this study, we used multiplex fluorophore imaging to track the fate of individual strands of HJ substrates. We report the novel finding that TRF2 inhibits WRN helicase strand displacement of HJs with telomeric repeats in duplex arms, but unwinding of HJs with a telomeric center or lacking telomeric sequence is unaffected. These data, together with results using TRF2 fragments and TRF2 HJ binding assays, indicate that both the TRF2 B- and Myb domains are required to inhibit WRN HJ activity. We propose a novel model whereby simultaneous binding of the TRF2 B-domain to the HJ core and the Myb domain to telomeric arms promote and stabilize HJs in a stacked arm conformation that is unfavorable for unwinding. Our biochemical study provides a mechanistic basis for the cellular findings that TRF2 regulates WRN activity at telomeres.
Figures







Similar articles
-
TRF2 recruits the Werner syndrome (WRN) exonuclease for processing of telomeric DNA.Oncogene. 2004 Jan 8;23(1):149-56. doi: 10.1038/sj.onc.1206906. Oncogene. 2004. PMID: 14712220
-
Sequence-specific processing of telomeric 3' overhangs by the Werner syndrome protein exonuclease activity.Aging (Albany NY). 2009 Mar 17;1(3):289-302. doi: 10.18632/aging.100032. Aging (Albany NY). 2009. PMID: 20157518 Free PMC article.
-
The Werner syndrome helicase and exonuclease cooperate to resolve telomeric D loops in a manner regulated by TRF1 and TRF2.Mol Cell. 2004 Jun 18;14(6):763-74. doi: 10.1016/j.molcel.2004.05.023. Mol Cell. 2004. PMID: 15200954
-
Telomere ResQue and preservation--roles for the Werner syndrome protein and other RecQ helicases.Mech Ageing Dev. 2008 Jan-Feb;129(1-2):79-90. doi: 10.1016/j.mad.2007.10.007. Epub 2007 Oct 30. Mech Ageing Dev. 2008. PMID: 18054793 Review.
-
Interstitial telomeric loops and implications of the interaction between TRF2 and lamin A/C.Differentiation. 2018 Jul-Aug;102:19-26. doi: 10.1016/j.diff.2018.06.001. Epub 2018 Jun 18. Differentiation. 2018. PMID: 29979997 Review.
Cited by
-
Telomeres and Cancer: Resolving the Paradox.Annu Rev Cancer Biol. 2021 Mar;5(1):59-77. doi: 10.1146/annurev-cancerbio-050420-023410. Annu Rev Cancer Biol. 2021. PMID: 34532611 Free PMC article.
-
Unraveling the roles of WRN and DNA-PKcs at telomeres.Aging (Albany NY). 2010 May;2(5):257-8. doi: 10.18632/aging.100150. Aging (Albany NY). 2010. PMID: 20519782 Free PMC article. No abstract available.
-
The telomere binding protein TRF2 induces chromatin compaction.PLoS One. 2011 Apr 19;6(4):e19124. doi: 10.1371/journal.pone.0019124. PLoS One. 2011. PMID: 21526145 Free PMC article.
-
The DNA structure and sequence preferences of WRN underlie its function in telomeric recombination events.Nat Commun. 2015 Sep 30;6:8331. doi: 10.1038/ncomms9331. Nat Commun. 2015. PMID: 26420422 Free PMC article.
-
Human Rap1 modulates TRF2 attraction to telomeric DNA.Nucleic Acids Res. 2015 Mar 11;43(5):2691-700. doi: 10.1093/nar/gkv097. Epub 2015 Feb 11. Nucleic Acids Res. 2015. PMID: 25675958 Free PMC article.
References
-
- Kudlow BA, Kennedy BK, Monnat RJ., Jr Werner and Hutchinson-Gilford progeria syndromes: mechanistic basis of human progeroid diseases. Nat. Rev. Mol. Cell Biol. 2007;8:394–404. - PubMed
-
- Opresko PL. Telomere ResQue and preservation–roles for the Werner syndrome protein and other RecQ helicases. Mech. Ageing Dev. 2008;129:79–90. - PubMed
-
- Chu WK, Hickson ID. RecQ helicases: multifunctional genome caretakers. Nat. Rev. Cancer. 2009;9:644–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
