

Dominant mutations in the late 40S biogenesis factor Ltv1 affect cytoplasmic maturation of the small ribosomal subunit in Saccharomyces cerevisiae
- PMID: 20215468
- PMCID: PMC2870955
- DOI: 10.1534/genetics.110.115584
Dominant mutations in the late 40S biogenesis factor Ltv1 affect cytoplasmic maturation of the small ribosomal subunit in Saccharomyces cerevisiae
Abstract
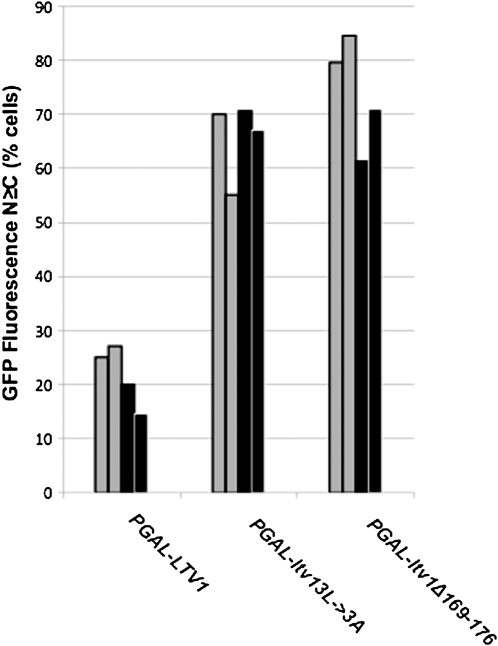
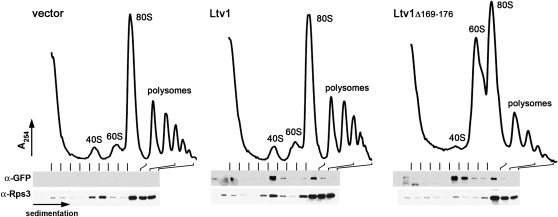
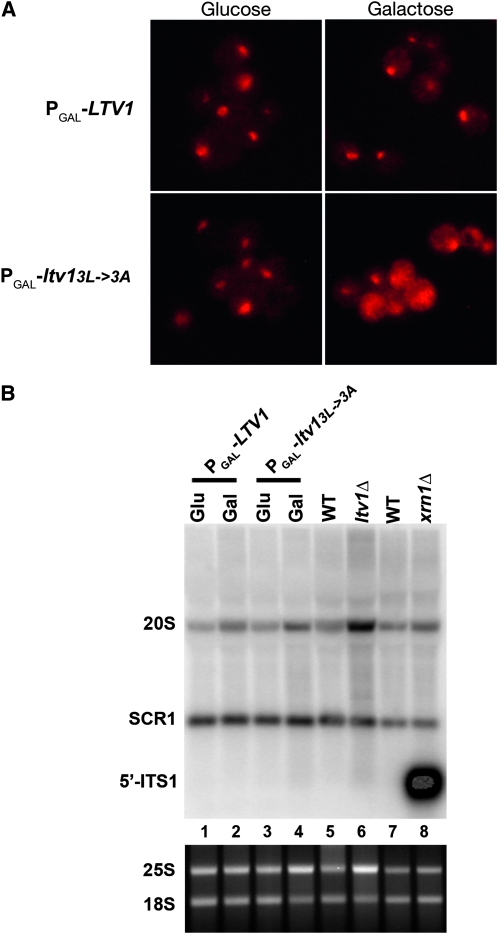
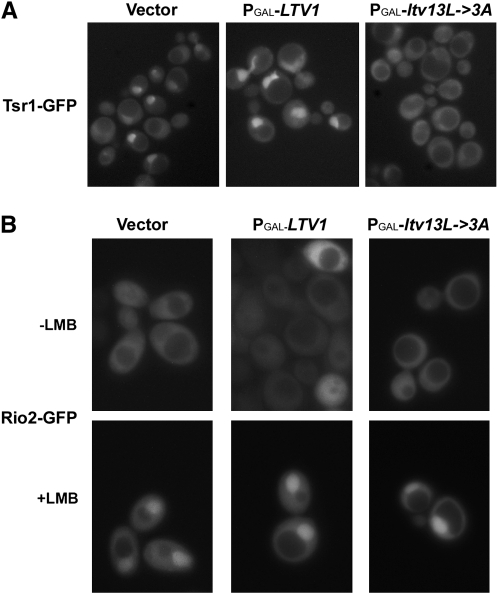
In eukaryotes, 40S and 60S ribosomal subunits are assembled in the nucleus from rRNAs and ribosomal proteins, exported as premature complexes, and processed in final maturation steps in the cytoplasm. Ltv1 is a conserved 40S ribosome biogenesis factor that interacts with pre-40S complexes in vivo and is proposed to function in yeast in nuclear export. Cells lacking LTV1 grow slowly and are significantly impaired in mature 40S subunit production. Here we show that mutation or deletion of a putative nuclear export sequence in LTV1 is strongly dominant negative, but the protein does not accumulate in the nucleus, as expected for a mutation affecting export. In fact, most of the mutant protein is cytoplasmic and associated with pre-40S subunits. Cells expressing mutant Ltv1 have a 40S biogenesis defect, accumulate 20S rRNA in the cytoplasm as detected by FISH, and retain the late-acting biogenesis factor Tsr1 in the cytoplasm. Finally, overexpression of mutant Ltv1 is associated with nuclear retention of 40S subunit marker proteins, RpS2-GFP and RpS3-GFP. We suggest that the proximal consequence of these LTV1 mutations is inhibition of the cytoplasmic maturation of 40S subunits and that nuclear retention of pre-40S subunits is a downstream consequence of the failure to release and recycle critical factors back to the nucleus.
Figures


Similar articles
-
Genetic analysis of the ribosome biogenesis factor Ltv1 of Saccharomyces cerevisiae.Genetics. 2014 Nov;198(3):1071-85. doi: 10.1534/genetics.114.168294. Epub 2014 Sep 10. Genetics. 2014. PMID: 25213169 Free PMC article.
-
Ltv1 is required for efficient nuclear export of the ribosomal small subunit in Saccharomyces cerevisiae.Genetics. 2006 Oct;174(2):679-91. doi: 10.1534/genetics.106.062117. Epub 2006 Aug 3. Genetics. 2006. PMID: 16888326 Free PMC article.
-
Role of the 40S beak ribosomal protein eS12 in ribosome biogenesis and function in Saccharomyces cerevisiae.RNA Biol. 2020 Sep;17(9):1261-1276. doi: 10.1080/15476286.2020.1767951. Epub 2020 Jun 7. RNA Biol. 2020. PMID: 32408794 Free PMC article.
-
Nuclear export and cytoplasmic maturation of ribosomal subunits.FEBS Lett. 2007 Jun 19;581(15):2783-93. doi: 10.1016/j.febslet.2007.05.013. Epub 2007 May 11. FEBS Lett. 2007. PMID: 17509569 Review.
-
Mechanistic insight into eukaryotic 60S ribosomal subunit biogenesis by cryo-electron microscopy.RNA. 2016 Nov;22(11):1643-1662. doi: 10.1261/rna.057927.116. RNA. 2016. PMID: 27875256 Free PMC article. Review.
Cited by
-
Role of Mex67-Mtr2 in the nuclear export of 40S pre-ribosomes.PLoS Genet. 2012;8(8):e1002915. doi: 10.1371/journal.pgen.1002915. Epub 2012 Aug 30. PLoS Genet. 2012. PMID: 22956913 Free PMC article.
-
Hairless Streaks in Cattle Implicate TSR2 in Early Hair Follicle Formation.PLoS Genet. 2015 Jul 23;11(7):e1005427. doi: 10.1371/journal.pgen.1005427. eCollection 2015 Jul. PLoS Genet. 2015. PMID: 26203908 Free PMC article.
-
Drosophila Low Temperature Viability Protein 1 (LTV1) Is Required for Ribosome Biogenesis and Cell Growth Downstream of Drosophila Myc (dMyc).J Biol Chem. 2015 May 22;290(21):13591-604. doi: 10.1074/jbc.M114.607036. Epub 2015 Apr 9. J Biol Chem. 2015. PMID: 25858587 Free PMC article.
-
The eukaryote-specific N-terminal extension of ribosomal protein S31 contributes to the assembly and function of 40S ribosomal subunits.Nucleic Acids Res. 2016 Sep 19;44(16):7777-91. doi: 10.1093/nar/gkw641. Epub 2016 Jul 15. Nucleic Acids Res. 2016. PMID: 27422873 Free PMC article.
-
Structural Heterogeneity in Pre-40S Ribosomes.Structure. 2017 Feb 7;25(2):329-340. doi: 10.1016/j.str.2016.12.011. Epub 2017 Jan 19. Structure. 2017. PMID: 28111018 Free PMC article.
References
-
- Amberg, D. C., A. L. Goldstein and C. N. Cole, 1992. Isolation and characterization of RAT1: an essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes Dev. 6 1173–1189. - PubMed
-
- Brand, R. C., J. Klootwijk, T. J. Van Steenbergen, A. J. De Kok and R. J. Planta, 1977. Secondary methylation of yeast ribosomal precursor RNA. Eur. J. Biochem. 75 311–318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
