

Antitumor activity of type I and type III interferons in BNL hepatoma model
- PMID: 20217081
- PMCID: PMC4699672
- DOI: 10.1007/s00262-010-0831-3
Antitumor activity of type I and type III interferons in BNL hepatoma model
Abstract
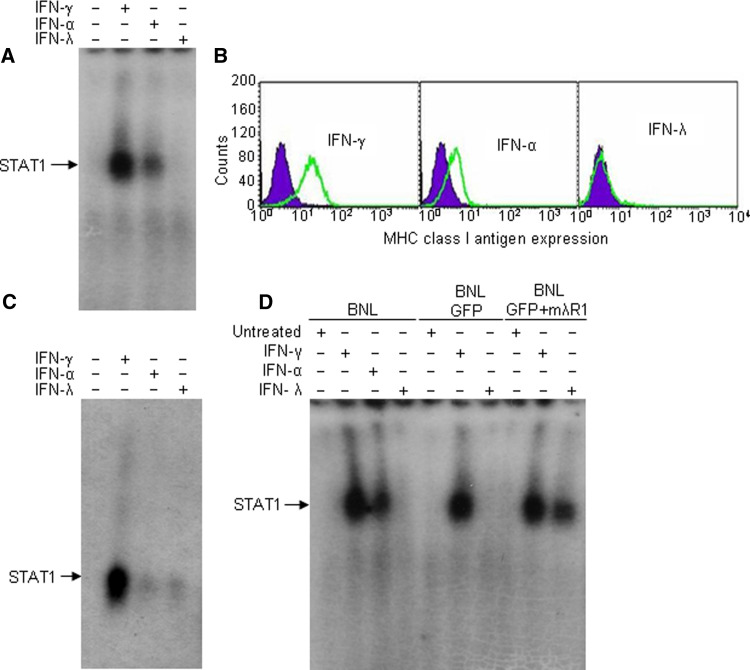
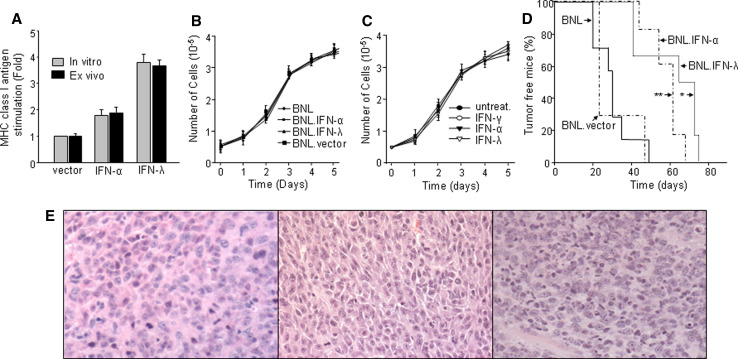
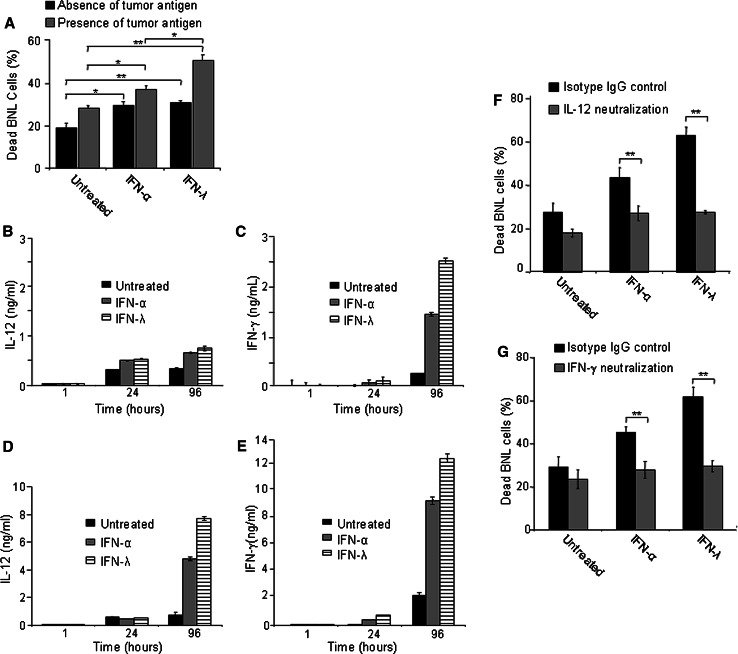
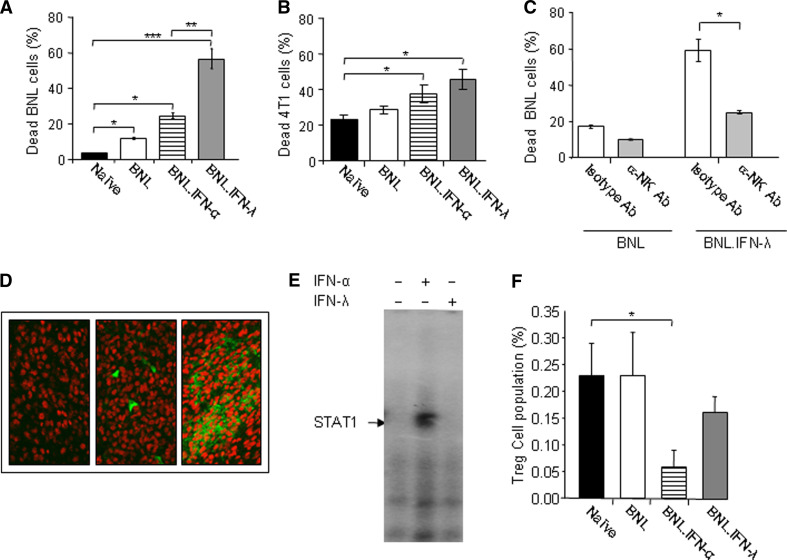
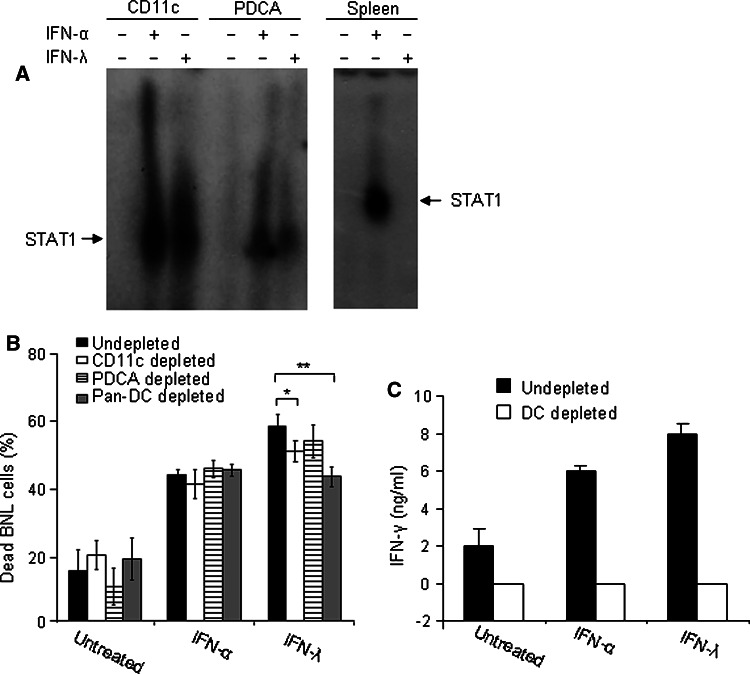
Hepatocellular carcinoma (HCC) occurs most commonly secondary to cirrhosis due to chronic hepatitis C or B virus (HCV/HBV) infections. Type I interferon (IFN-alpha) treatment of chronic HCV/HBV infections reduces the incidence of HCC in cirrhotic patients. However, IFN-alpha toxicity limits its tolerability and efficacy highlighting a need for better therapeutic treatments. A recently discovered type III IFN (IFN-lambda) has been shown to possess antiviral properties against HCV and HBV in vitro. In phase I clinical trials, IFN-lambda treatment did not cause significant adverse reactions. Using a gene therapy approach, we compared the antitumor properties of IFN-alpha and IFN-lambda in a transplantable hepatoma model of HCC. BALB/c mice were inoculated with syngeneic BNL hepatoma cells, or BNL cells expressing IFN-lambda (BNL.IFN-lambda cells) or IFN-alpha (BNL.IFN-alpha cells). Despite the lack of antiproliferative activity of IFNs on BNL cells, both BNL.IFN-lambda and BNL.IFN-alpha cells displayed retarded growth kinetics in vivo. Depletion of NK cells from splenocytes inhibited splenocyte-mediated cytotoxicity, demonstrating that NK cells play a role in IFN-induced antitumor responses. However, isolated NK cells did not respond directly to IFN-lambda. There was also a marked NK cell infiltration in IFN-lambda producing tumors. In addition, IFN-lambda and, to a lesser extent, IFN-alpha enhanced immunocytotoxicity of splenocytes primed with irradiated BNL cells. Splenocyte cytotoxicity against BNL cells was dependent on IL-12 and IFN-gamma, and mediated by dendritic cells. In contrast to NK cells, isolated from spleen CD11c+ and mPDCA+ dendritic cells responded directly to IFN-lambda. The antitumor activities of IFN-lambda against hepatoma, in combination with HCV and HBV antiviral activities warrant further investigation into the clinical use of IFN-lambda to prevent HCC in HCV/HBV-infected cirrhotic patients, as well as to treat liver cancer.
Figures
References
-
- Lau WY, Lai EC. Hepatocellular carcinoma: current management and recent advances. Hepatobiliary Pancreat Dis Int. 2008;7:237–257. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
