

The neonatal Fc receptor as therapeutic target in IgG-mediated autoimmune diseases
- PMID: 20217455
- PMCID: PMC11115620
- DOI: 10.1007/s00018-010-0318-6
The neonatal Fc receptor as therapeutic target in IgG-mediated autoimmune diseases
Abstract
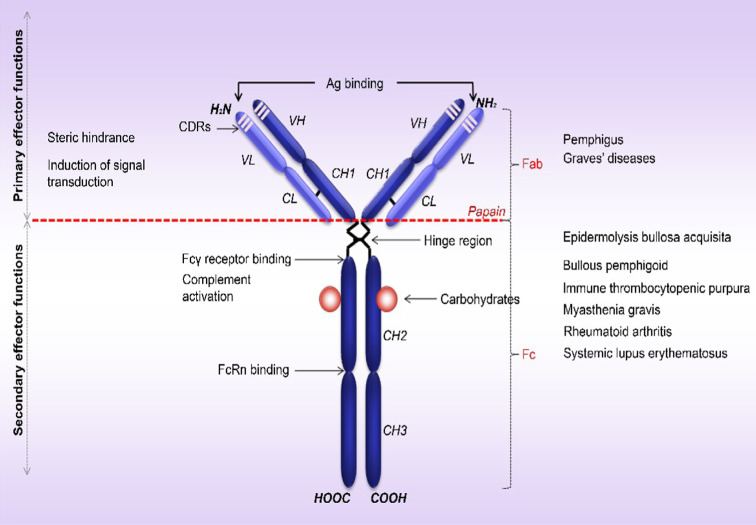
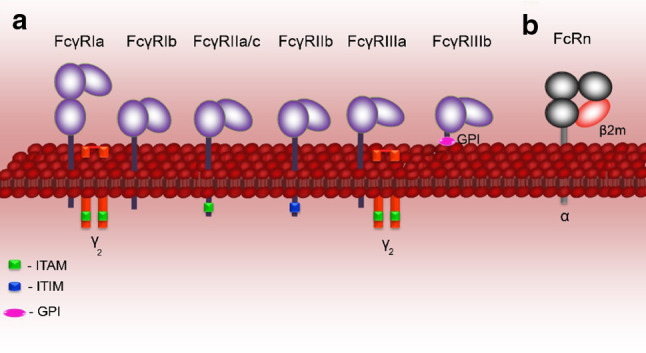
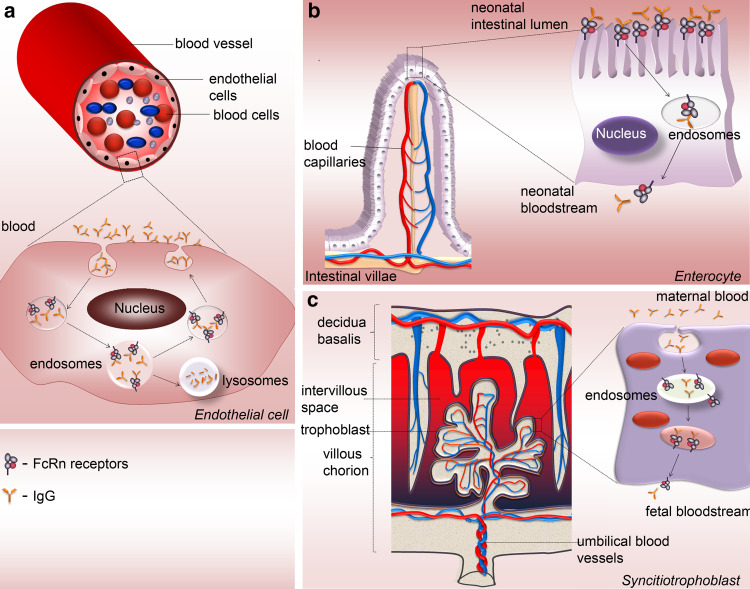
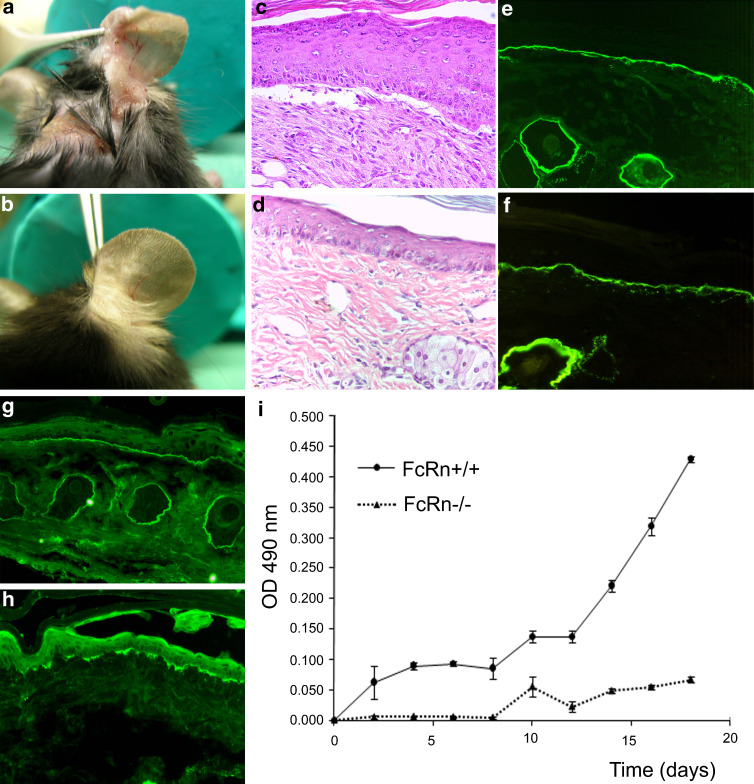
Therapy approaches based on lowering levels of pathogenic autoantibodies represent rational, effective, and safe treatment modalities of autoimmune diseases. The neonatal Fc receptor (FcRn) is a major factor regulating the serum levels of IgG antibodies. While FcRn-mediated half-life extension is beneficial for IgG antibody responses against pathogens, it also prolongs the serum half-life of IgG autoantibodies and thus promotes tissue damage in autoimmune diseases. In the present review article, we examine current evidence on the relevance of FcRn in maintaining high autoantibody levels and discuss FcRn-targeted therapeutic approaches. Further investigation of the FcRn-IgG interaction will not only provide mechanistic insights into the receptor function, but should also greatly facilitate the design of therapeutics combining optimal pharmacokinetic properties with the appropriate antibody effector functions in autoimmune diseases.
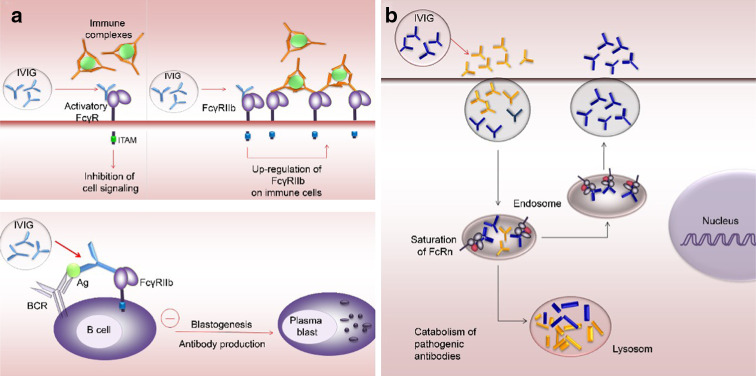
Figures
References
-
- Sinha AA, Lopez MT, McDevitt HO. Autoimmune diseases: the failure of self tolerance. Science. 1990;248:1380–1388. - PubMed
-
- Bach J. The effect of infections on susceptibility to autoimmune and allergic diseases. N Engl J Med. 2002;347:911–920. - PubMed
-
- Drachman DB. How to recognize an antibody-mediated autoimmune disease: criteria. Res Publ Assoc Res Nerv Ment Dis. 1990;68:183–186. - PubMed
-
- Rose NR, Bona C. Defining criteria for autoimmune diseases (Witebsky’s postulates revisited) Immunol Today. 1993;14:426–430. - PubMed
-
- Stassi G, De Maria R. Autoimmune thyroid disease: new models of cell death in autoimmunity. Nat Rev Immunol. 2002;2:195–204. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
