

Translation control by protein kinase R restricts minute virus of mice infection: role in parvovirus oncolysis
- PMID: 20219905
- PMCID: PMC2863853
- DOI: 10.1128/JVI.02188-09
Translation control by protein kinase R restricts minute virus of mice infection: role in parvovirus oncolysis
Abstract
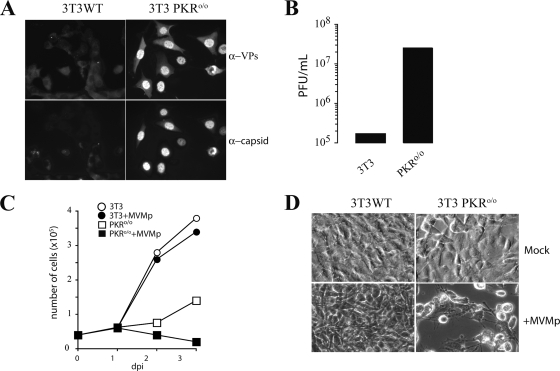
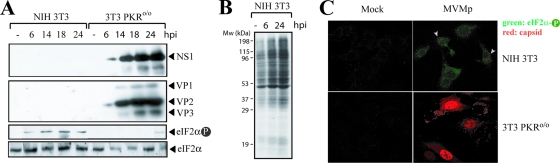
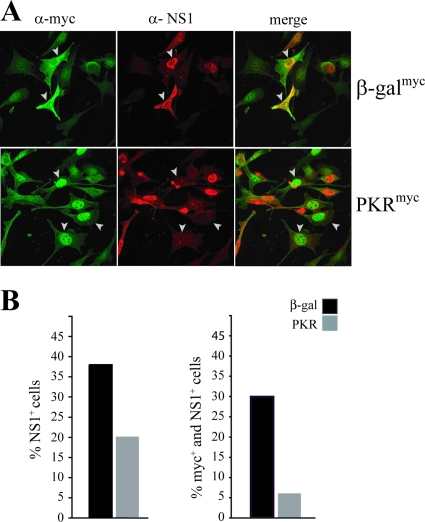
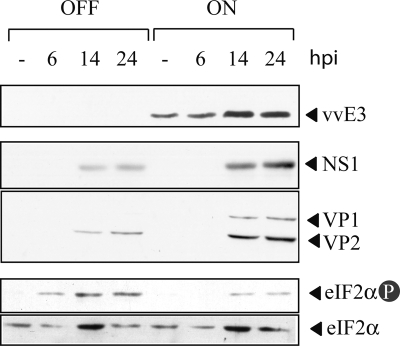
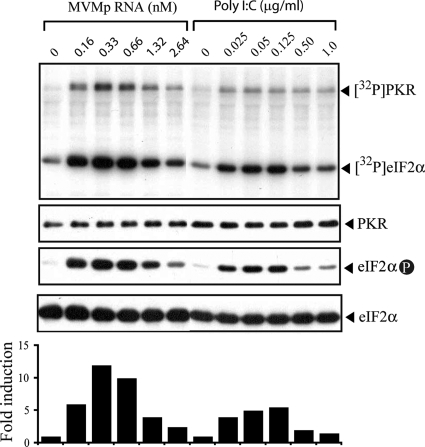
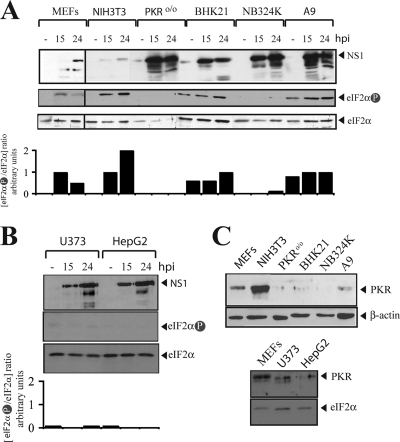
The relevance of translational control in the gene expression and oncotropism of the autonomous parvoviruses was investigated with MVMp, the prototype strain of minute virus of mice (MVM), infecting normal and transformed rodent and human cells of different tissue origins. Mouse embryo fibroblasts (MEFs) and NIH 3T3 fibroblasts were resistant to MVMp infection, but 3T3 fibroblasts derived from double-stranded RNA (dsRNA)-dependent protein kinase R (PKR) knockout mice (PKR(o/o)) behaved in a manner that was highly permissive to productive MVMp replication. NIH 3T3 resistance correlated with significant phosphorylation of eukaryotic translation initiation factor 2 (eIF2) occurring at early time points after infection. Permissive PKR(o/o) cells were converted to MVMp-restrictive cells after reintroduction of the PKR gene by transfection. Conversely, regulated expression of the vaccinia virus E3 protein, a PKR inhibitor, in MEFs prevented eIF2alpha phosphorylation and increased MVMp protein synthesis. In vitro-synthesized genome-length R1 mRNA of MVMp was a potent activator of PKR. Virus-resistant primary MEFs and NIH 3T3 cells responded to MVMp infection with significant increases in eIF2alpha phosphorylation. In contrast, virus-permissive mouse (PKR(o/o), BHK21, and A9) and human transformed (NB324K fibroblast, U373 glioma, and HepG2 hepatoma) cells consistently showed no significant increase in the level of eIF2alpha phosphorylation following MVMp infection. The synthesis of the viral NS1 protein was inversely correlated with the steady-state PKR levels. Our results show that the PKR-mediated antiviral response is an important mechanism for control of productive MVMp infection, and its impairment in human transformed cells allowed efficient MVMp gene expression. PKR translational control may therefore contribute to the oncolysis of MVMp and other autonomous parvoviruses.
Figures
Similar articles
-
Translational resistance of late alphavirus mRNA to eIF2alpha phosphorylation: a strategy to overcome the antiviral effect of protein kinase PKR.Genes Dev. 2006 Jan 1;20(1):87-100. doi: 10.1101/gad.357006. Genes Dev. 2006. PMID: 16391235 Free PMC article.
-
Viral host-range factor C7 or K1 is essential for modified vaccinia virus Ankara late gene expression in human and murine cells, irrespective of their capacity to inhibit protein kinase R-mediated phosphorylation of eukaryotic translation initiation factor 2alpha.J Gen Virol. 2010 Feb;91(Pt 2):470-82. doi: 10.1099/vir.0.015347-0. Epub 2009 Oct 21. J Gen Virol. 2010. PMID: 19846675
-
The double-stranded RNA-activated protein kinase PKR is dispensable for regulation of translation initiation in response to either calcium mobilization from the endoplasmic reticulum or essential amino acid starvation.Biochem Biophys Res Commun. 2001 Jan 12;280(1):293-300. doi: 10.1006/bbrc.2000.4103. Biochem Biophys Res Commun. 2001. PMID: 11162513
-
HIV-1 translation and its regulation by cellular factors PKR and PACT.Virus Res. 2014 Nov 26;193:65-77. doi: 10.1016/j.virusres.2014.07.014. Epub 2014 Jul 23. Virus Res. 2014. PMID: 25064266 Review.
-
The interferon system and vaccinia virus evasion mechanisms.J Interferon Cytokine Res. 2009 Sep;29(9):581-98. doi: 10.1089/jir.2009.0073. J Interferon Cytokine Res. 2009. PMID: 19708815 Review.
Cited by
-
GCN2 has inhibitory effect on human immunodeficiency virus-1 protein synthesis and is cleaved upon viral infection.PLoS One. 2012;7(10):e47272. doi: 10.1371/journal.pone.0047272. Epub 2012 Oct 23. PLoS One. 2012. PMID: 23110064 Free PMC article.
-
Adaptive changes in alphavirus mRNA translation allowed colonization of vertebrate hosts.J Virol. 2012 Sep;86(17):9484-94. doi: 10.1128/JVI.01114-12. Epub 2012 Jul 3. J Virol. 2012. PMID: 22761388 Free PMC article.
-
Complementary induction of immunogenic cell death by oncolytic parvovirus H-1PV and gemcitabine in pancreatic cancer.J Virol. 2014 May;88(10):5263-76. doi: 10.1128/JVI.03688-13. Epub 2014 Feb 26. J Virol. 2014. PMID: 24574398 Free PMC article.
-
Oncolytic Rodent Protoparvoviruses Evade a TLR- and RLR-Independent Antiviral Response in Transformed Cells.Pathogens. 2023 Apr 17;12(4):607. doi: 10.3390/pathogens12040607. Pathogens. 2023. PMID: 37111493 Free PMC article.
-
Oncolytic virus therapy for glioblastoma multiforme: concepts and candidates.Cancer J. 2012 Jan-Feb;18(1):69-81. doi: 10.1097/PPO.0b013e31824671c9. Cancer J. 2012. PMID: 22290260 Free PMC article. Review.
References
-
- Angelova, A. L., M. Aprahamian, G. Balboni, H. J. Delecluse, R. Feederle, I. Kiprianova, S. P. Grekova, A. S. Galabov, M. Witzens-Harig, A. D. Ho, J. Rommelaere, and Z. Raykov. 2009. Oncolytic rat parvovirus H-1PV, a candidate for the treatment of human lymphoma: In vitro and in vivo studies. Mol. Ther. 17:1164-1172. - PMC - PubMed
-
- Balachandran, S., and G. N. Barber. 2004. Defective translational control facilitates vesicular stomatitis virus oncolysis. Cancer Cell 5:51-65. - PubMed
-
- Balachandran, S., P. C. Roberts, L. E. Brown, H. Truong, A. K. Pattnaik, D. R. Archer, and G. N. Barber. 2000. Essential role for the dsRNA-dependent protein kinase PKR in innate immunity to viral infection. Immunity 13:129-141. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
