

Bidirectional regulation of hippocampal long-term synaptic plasticity and its influence on opposing forms of memory
- PMID: 20220016
- PMCID: PMC6632240
- DOI: 10.1523/JNEUROSCI.1330-09.2010
Bidirectional regulation of hippocampal long-term synaptic plasticity and its influence on opposing forms of memory
Abstract
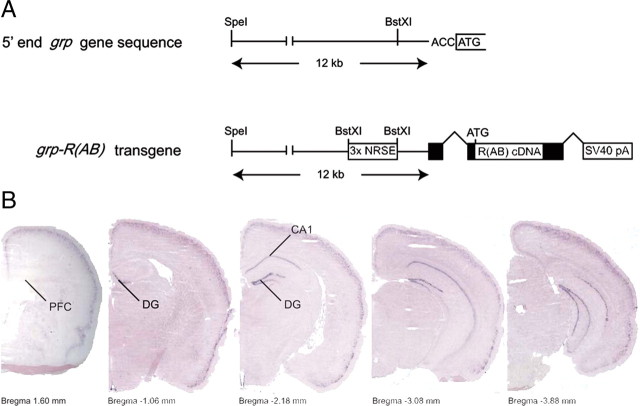
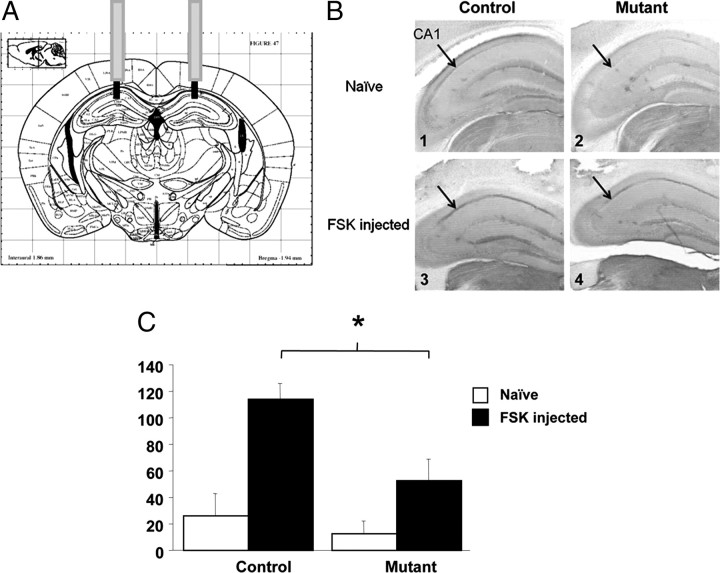
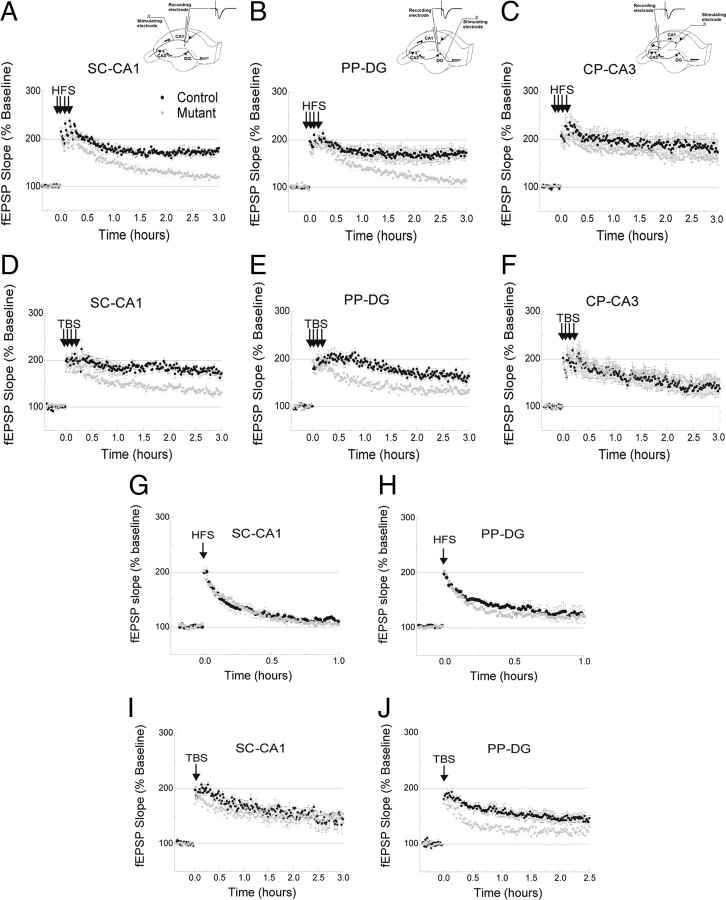
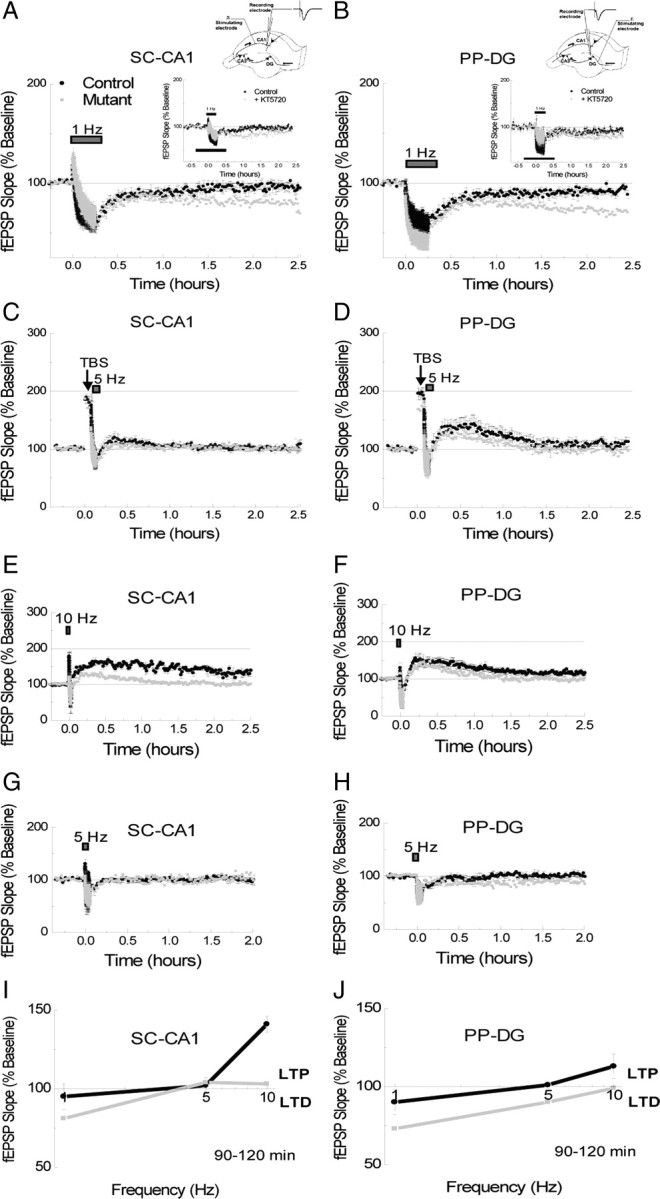
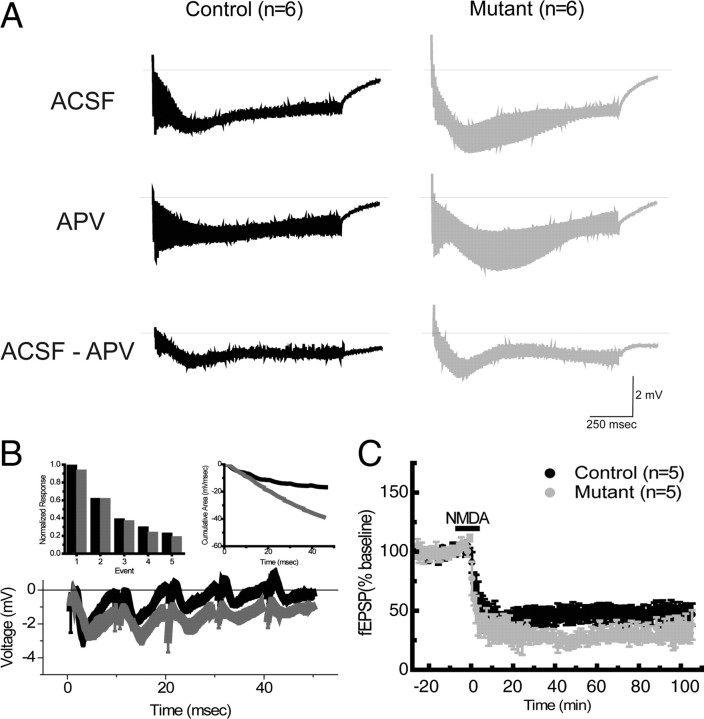
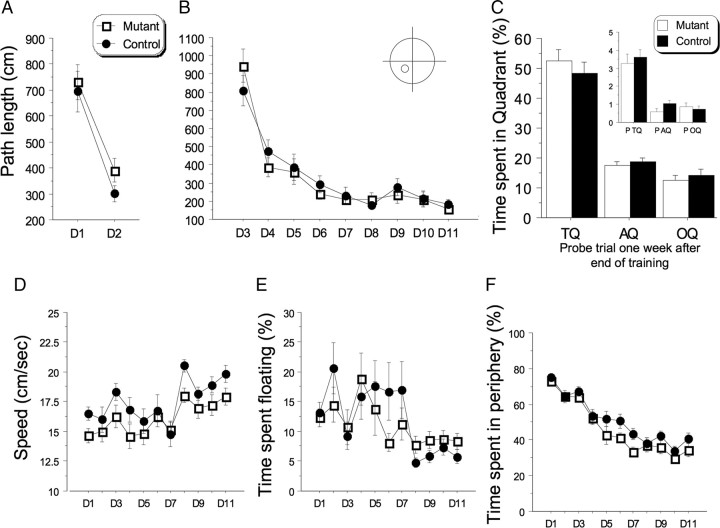
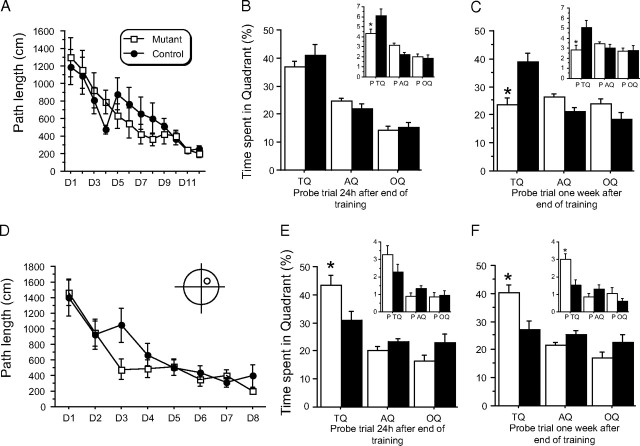
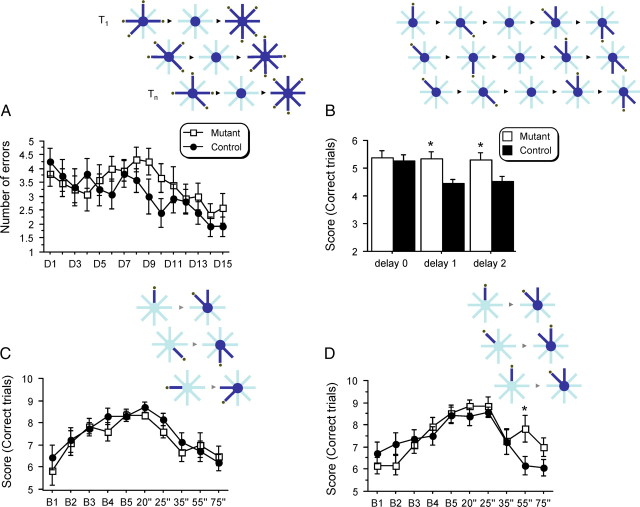
Reference memory characterizes the long-term storage of information acquired through numerous trials. In contrast, working memory represents the short-term acquisition of trial-unique information. A number of studies in the rodent hippocampus have focused on the contribution of long-term synaptic potentiation (LTP) to long-term reference memory. In contrast, little is known about the synaptic plasticity correlates of hippocampal-based components of working memory. Here, we described a mouse with selective expression of a dominant-negative mutant of the regulatory subunit of protein kinase A (PKA) only in two regions of the hippocampus, the dentate gyrus and area CA1. This mouse showed a deficit in several forms of LTP in both hippocampal subregions and a lowered threshold for the consolidation of long-term synaptic depression (LTD). When trained with one trial per day in a water maze task, mutant mice displayed a deficit in consolidation of long-term memory. In contrast, these mice proved to be more flexible after a transfer test and also showed a delay-dependent increased performance in working memory, when repetitive information (proactive interference) was presented. We suggest that through its bidirectional control over synaptic plasticity PKA can regulate opposing forms of memory. The defect in L-LTP disrupts long-term memory consolidation. The persistence of LTD may allow acquisition of new information by restricting the body of previously stored information and suppressing interference.
Figures
Similar articles
-
Regulation of hippocampal synaptic plasticity by cyclic AMP-dependent protein kinases.Prog Neurobiol. 2003 Dec;71(6):401-37. doi: 10.1016/j.pneurobio.2003.12.003. Prog Neurobiol. 2003. PMID: 15013227 Review.
-
Genetic deletion of melanin-concentrating hormone neurons impairs hippocampal short-term synaptic plasticity and hippocampal-dependent forms of short-term memory.Hippocampus. 2015 Nov;25(11):1361-73. doi: 10.1002/hipo.22442. Epub 2015 Apr 2. Hippocampus. 2015. PMID: 25808129
-
Differential maintenance and frequency-dependent tuning of LTP at hippocampal synapses of specific strains of inbred mice.J Neurophysiol. 2000 Nov;84(5):2484-93. doi: 10.1152/jn.2000.84.5.2484. J Neurophysiol. 2000. PMID: 11067991
-
Environmental enrichment modifies the PKA-dependence of hippocampal LTP and improves hippocampus-dependent memory.Learn Mem. 2001 Jan-Feb;8(1):26-34. doi: 10.1101/lm.36301. Learn Mem. 2001. PMID: 11160761 Free PMC article.
-
LTD, LTP, and the sliding threshold for long-term synaptic plasticity.Hippocampus. 1996;6(1):35-42. doi: 10.1002/(SICI)1098-1063(1996)6:1<35::AID-HIPO7>3.0.CO;2-6. Hippocampus. 1996. PMID: 8878740 Review.
Cited by
-
Levels of Interference in Long and Short-Term Memory Differentially Modulate Non-REM and REM Sleep.Sleep. 2016 Dec 1;39(12):2173-2188. doi: 10.5665/sleep.6322. Sleep. 2016. PMID: 27748246 Free PMC article.
-
Differential changes in hippocampal CaMKII and GluA1 activity after memory training involving different levels of adaptive forgetting.Learn Mem. 2017 Jan 17;24(2):86-94. doi: 10.1101/lm.043505.116. Print 2017 Feb. Learn Mem. 2017. PMID: 28096498 Free PMC article.
-
Long-term effects of interference on short-term memory performance in the rat.PLoS One. 2017 Mar 13;12(3):e0173834. doi: 10.1371/journal.pone.0173834. eCollection 2017. PLoS One. 2017. PMID: 28288205 Free PMC article.
-
Weakened Intracellular Zn2+-Buffering in the Aged Dentate Gyrus and Its Involvement in Erasure of Maintained LTP.Mol Neurobiol. 2018 May;55(5):3856-3865. doi: 10.1007/s12035-017-0615-2. Epub 2017 May 25. Mol Neurobiol. 2018. PMID: 28547527
-
Differences in Synaptic Dysfunction Between rTg4510 and APP/PS1 Mouse Models of Alzheimer's Disease.J Alzheimers Dis. 2018;61(1):195-208. doi: 10.3233/JAD-170457. J Alzheimers Dis. 2018. PMID: 29154272 Free PMC article.
References
-
- Abel T, Nguyen PV, Barad M, Deuel TA, Kandel ER, Bourtchouladze R. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell. 1997;88:615–626. - PubMed
-
- Alarcón JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER, Barco A. Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron. 2004;42:947–959. - PubMed
-
- Arai A, Black J, Lynch G. Origins of the variations in long-term potentiation between synapses in the basal versus apical dendrites of hippocampal neurons. Hippocampus. 1994;4:1–9. - PubMed
-
- Arnsten AF, Ramos BP, Birnbaum SG, Taylor JR. Protein kinase A as a therapeutic target for memory disorders: rationale and challenges. Trends Mol Med. 2005;11:121–128. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous