

Independent epileptiform discharge patterns in the olfactory and limbic areas of the in vitro isolated Guinea pig brain during 4-aminopyridine treatment
- PMID: 20220076
- PMCID: PMC4878894
- DOI: 10.1152/jn.00862.2009
Independent epileptiform discharge patterns in the olfactory and limbic areas of the in vitro isolated Guinea pig brain during 4-aminopyridine treatment
Abstract
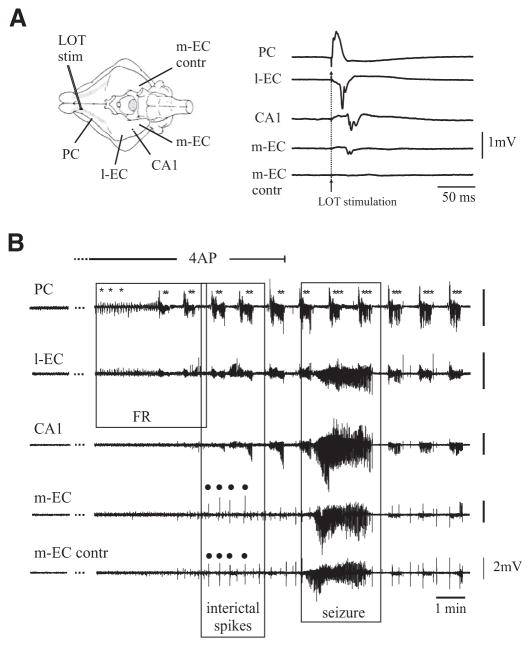
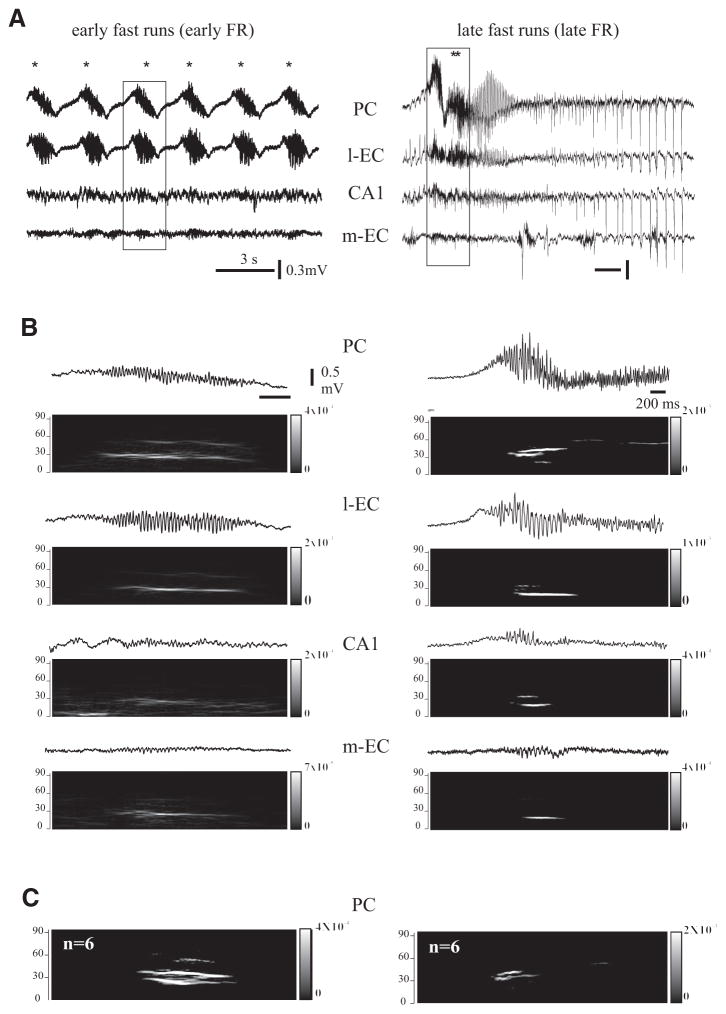
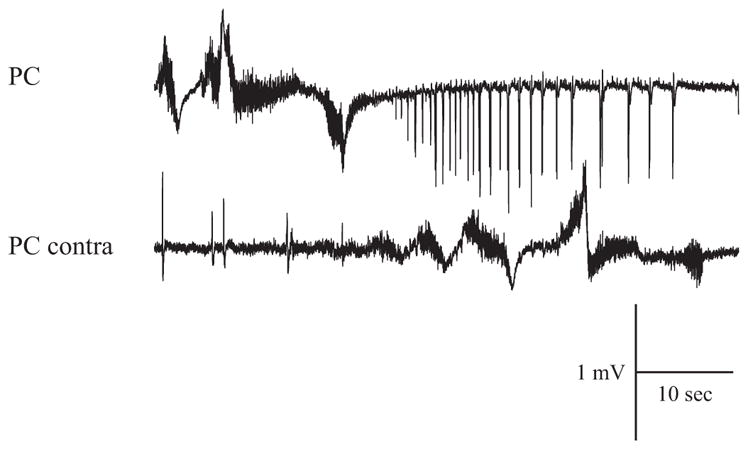
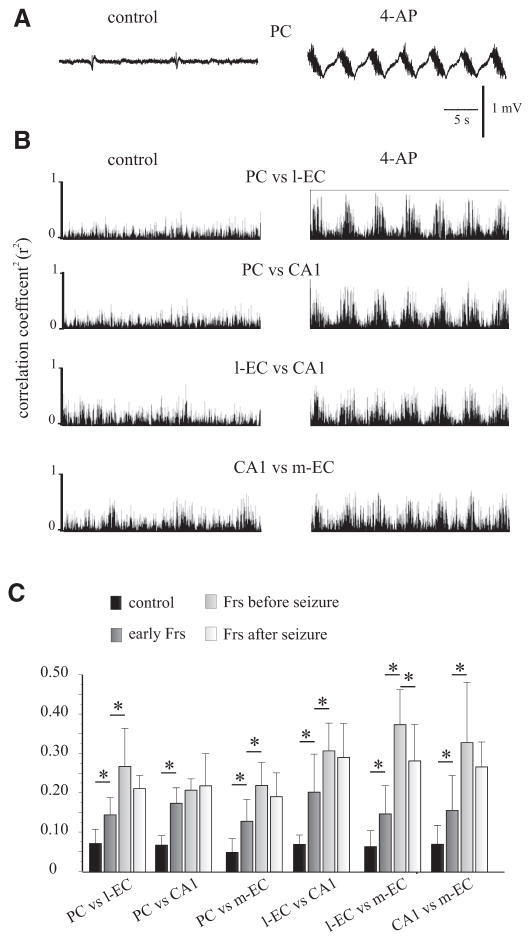
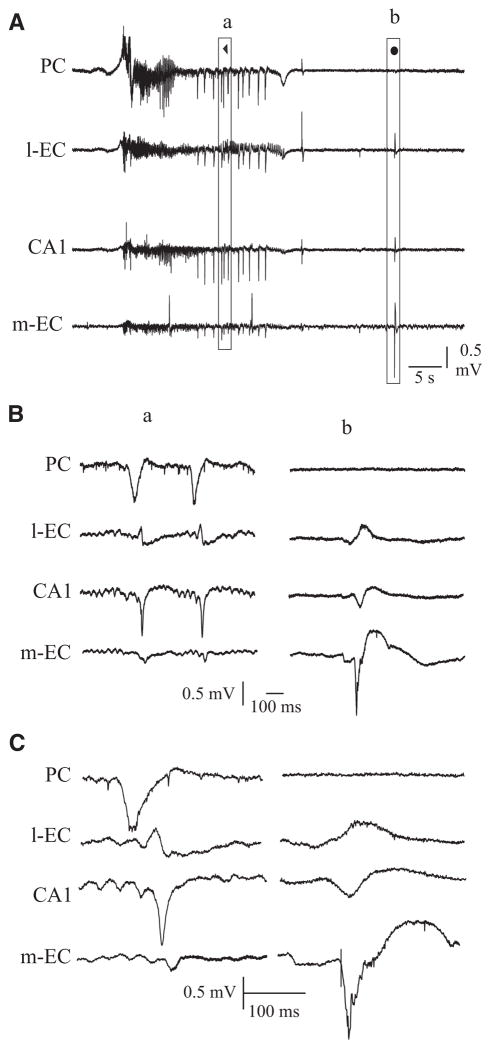
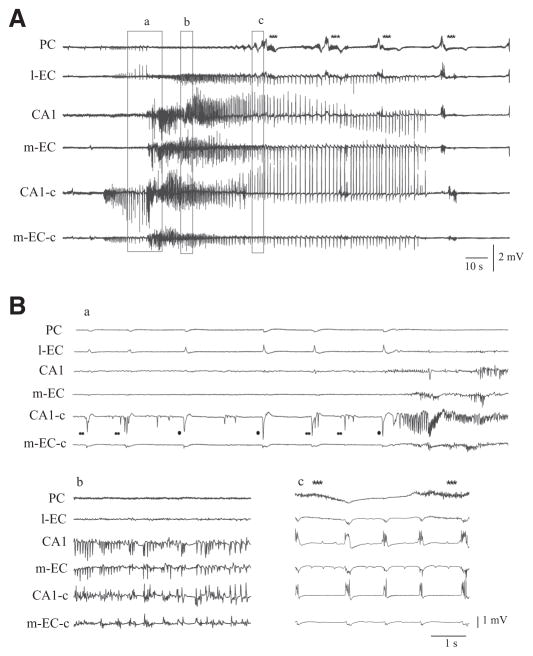
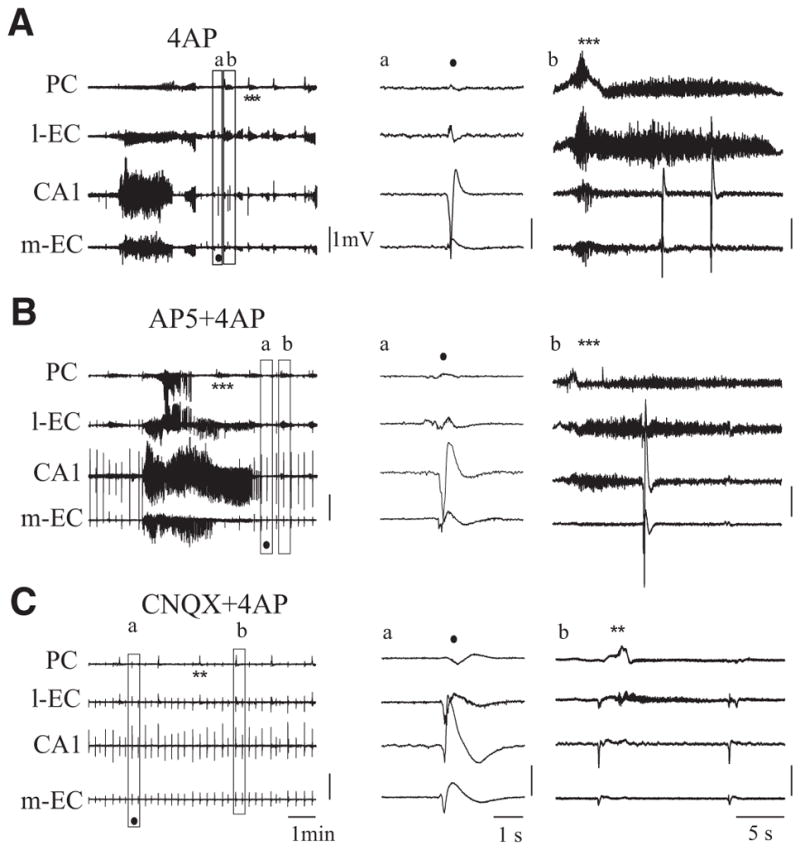
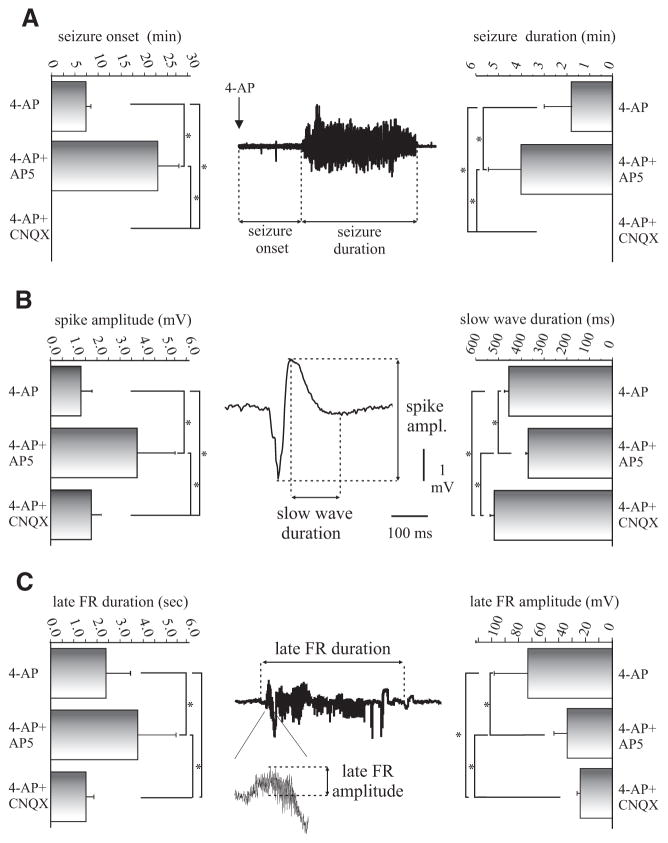
In vitro studies performed on brain slices demonstrate that the potassium channel blocker 4-aminopyridine (4AP, 50 microM) discloses electrographic seizure activity and interictal discharges. These epileptiform patterns have been further analyzed here in a isolated whole guinea pig brain in vitro by using field potential recordings in olfactory and limbic structures. In 8 of 13 experiments runs of fast oscillatory activity (fast runs, FRs) in the piriform cortex (PC) propagated to the lateral entorhinal cortex (EC), hippocampus and occasionally to the medial EC. Early and late FRs were asynchronous in the hemispheres showed different duration [1.78 +/- 0.51 and 27.95 +/- 4.55 (SD) s, respectively], frequency of occurrence (1.82 +/- 0.49 and 34.16 +/- 6.03 s) and frequency content (20-40 vs. 40-60 Hz). Preictal spikes independent from the FRs appeared in the hippocampus/EC and developed into ictal-like discharges that did not propagate to the PC. Ictal-like activity consisted of fast activity with onset either in the hippocampus (n = 6) or in the mEC (n = 2), followed by irregular spiking and sequences of diffusely synchronous bursts. Perfusion of the N-methyl-d-aspartate receptor antagonist 2-amino-5-phosphonopentanoic acid (100 microM) did not prevent FRs, increased the duration of limbic ictal-like discharges and favored their propagation to olfactory structures. The AMPA receptor antagonist 6,7-dinitroquinoxaline-2,3-dione (50 microM) blocked ictal-like events and reduced FRs. In conclusion, 4AP-induced epileptiform activities are asynchronous and independent in olfactory and hippocampal-entorhinal regions. Epileptiform discharges in the isolated guinea pig brain show different pharmacological properties compared with rodent in vitro slices.
Figures
Similar articles
-
Hippocampus-entorhinal cortex loop and seizure generation in the young rodent limbic system.J Neurophysiol. 2000 May;83(5):3183-7. doi: 10.1152/jn.2000.83.5.3183. J Neurophysiol. 2000. PMID: 10805716
-
Propagation dynamics of epileptiform activity acutely induced by bicuculline in the hippocampal-parahippocampal region of the isolated Guinea pig brain.Epilepsia. 2005 Dec;46(12):1914-25. doi: 10.1111/j.1528-1167.2005.00342.x. Epilepsia. 2005. PMID: 16393157
-
Synchronous GABA-mediated potentials and epileptiform discharges in the rat limbic system in vitro.J Neurosci. 1996 Jun 15;16(12):3912-24. doi: 10.1523/JNEUROSCI.16-12-03912.1996. J Neurosci. 1996. PMID: 8656285 Free PMC article.
-
Network and pharmacological mechanisms leading to epileptiform synchronization in the limbic system in vitro.Prog Neurobiol. 2002 Oct;68(3):167-207. doi: 10.1016/s0301-0082(02)00077-1. Prog Neurobiol. 2002. PMID: 12450487 Review.
-
Do interictal discharges promote or control seizures? Experimental evidence from an in vitro model of epileptiform discharge.Epilepsia. 2001;42 Suppl 3:2-4. doi: 10.1046/j.1528-1157.2001.042suppl.3002.x. Epilepsia. 2001. PMID: 11520313 Review.
Cited by
-
The piriform, perirhinal, and entorhinal cortex in seizure generation.Front Neural Circuits. 2015 May 29;9:27. doi: 10.3389/fncir.2015.00027. eCollection 2015. Front Neural Circuits. 2015. PMID: 26074779 Free PMC article. Review.
-
Initiation, Propagation, and Termination of Partial (Focal) Seizures.Cold Spring Harb Perspect Med. 2015 Jul 1;5(7):a022368. doi: 10.1101/cshperspect.a022368. Cold Spring Harb Perspect Med. 2015. PMID: 26134843 Free PMC article. Review.
-
Seizure-like discharges induced by 4-aminopyridine in the olfactory system of the in vitro isolated guinea pig brain.Epilepsia. 2013 Apr;54(4):605-15. doi: 10.1111/epi.12133. Epub 2013 Mar 18. Epilepsia. 2013. PMID: 23505998 Free PMC article.
-
Epileptiform synchronization and high-frequency oscillations in brain slices comprising piriform and entorhinal cortices.Neuroscience. 2014 Dec 5;281:258-68. doi: 10.1016/j.neuroscience.2014.09.065. Epub 2014 Oct 5. Neuroscience. 2014. PMID: 25290016 Free PMC article.
-
Persistent ictal-like activity in rat entorhinal/perirhinal cortex following washout of 4-aminopyridine.Epilepsy Res. 2011 May;94(3):163-76. doi: 10.1016/j.eplepsyres.2011.01.017. Epub 2011 Feb 25. Epilepsy Res. 2011. PMID: 21353480 Free PMC article.
References
-
- Aram JA, Michelson HB, Wong RKS. Synchronized GABAergic IPSPs recorded in the neocortex after blockade of synaptic transmission mediated by excitatory amino acids. J Neurophysiol. 1991;65:1034–1041. - PubMed
-
- Avoli M. GABA-mediated synchronous potentials and seizure generation. Epilepsia. 1996;37:1035–1042. - PubMed
-
- Avoli M, D’Antuono M, Louvel J, Kohling R, Biagini G, Pumain R, D’Arcangelo G, Tancredi V. Network pharmachological mechanisms leading to epileptiform synchronization in the limbic system in vitro. Progr Neurobiol. 2002;68:167–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical