

Mutant huntingtin fragments form oligomers in a polyglutamine length-dependent manner in vitro and in vivo
- PMID: 20220138
- PMCID: PMC2863238
- DOI: 10.1074/jbc.M109.093708
Mutant huntingtin fragments form oligomers in a polyglutamine length-dependent manner in vitro and in vivo
Abstract
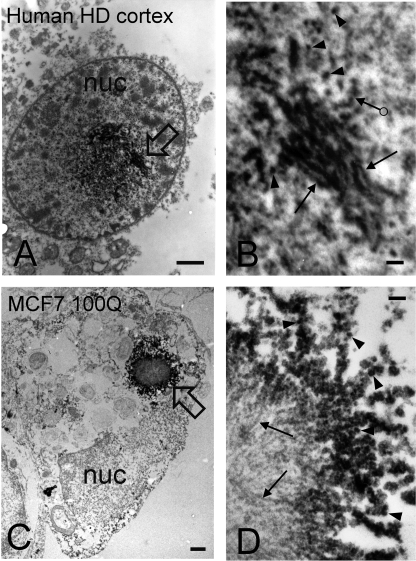
Huntington disease (HD) is caused by an expansion of more than 35-40 polyglutamine (polyQ) repeats in the huntingtin (htt) protein, resulting in accumulation of inclusion bodies containing fibrillar deposits of mutant htt fragments. Intriguingly, polyQ length is directly proportional to the propensity for htt to form fibrils and the severity of HD and is inversely correlated with age of onset. Although the structural basis for htt toxicity is unclear, the formation, abundance, and/or persistence of toxic conformers mediating neuronal dysfunction and degeneration in HD must also depend on polyQ length. Here we used atomic force microscopy to demonstrate mutant htt fragments and synthetic polyQ peptides form oligomers in a polyQ length-dependent manner. By time-lapse atomic force microscopy, oligomers form before fibrils, are transient in nature, and are occasionally direct precursors to fibrils. However, the vast majority of fibrils appear to form by monomer addition coinciding with the disappearance of oligomers. Thus, oligomers must undergo a major structural transition preceding fibril formation. In an immortalized striatal cell line and in brain homogenates from a mouse model of HD, a mutant htt fragment formed oligomers in a polyQ length-dependent manner that were similar in size to those formed in vitro, although these structures accumulated over time in vivo. Finally, using immunoelectron microscopy, we detected oligomeric-like structures in human HD brains. These results demonstrate that oligomer formation by a mutant htt fragment is strongly polyQ length-dependent in vitro and in vivo, consistent with a causative role for these structures, or subsets of these structures, in HD pathogenesis.
Figures
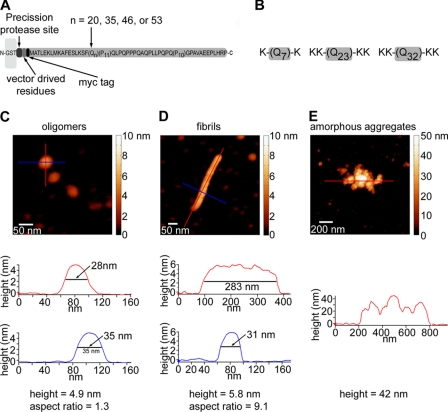
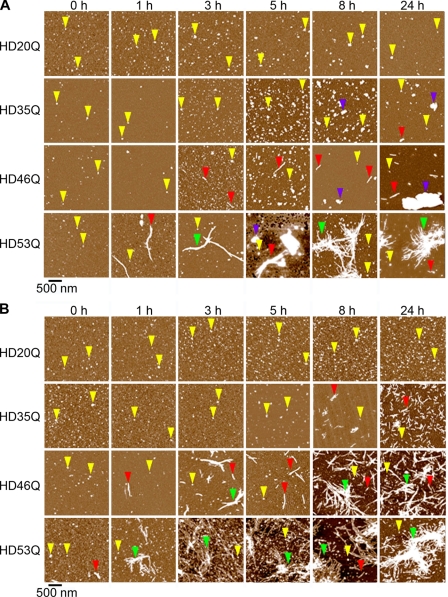
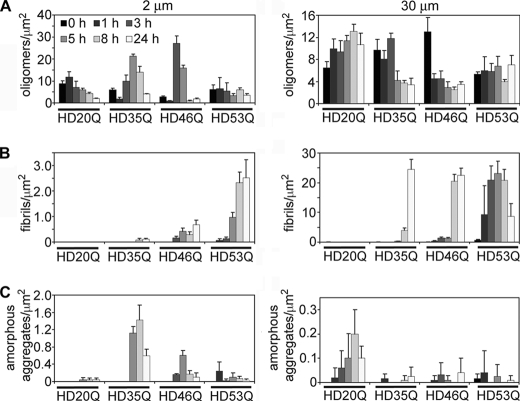

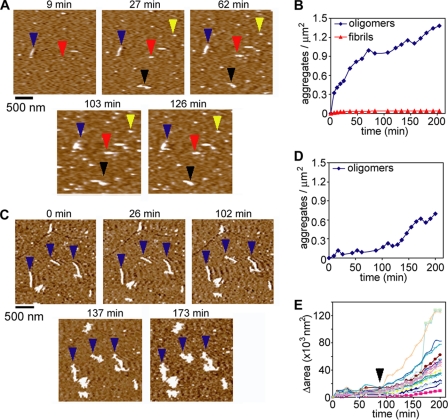



References
-
- The Huntington's Disease Collaborative Research Group (1993) Cell 72, 971–983 - PubMed
-
- DiFiglia M., Sapp E., Chase K. O., Davies S. W., Bates G. P., Vonsattel J. P., Aronin N. (1997) Science 277, 1990–1993 - PubMed
-
- Zoghbi H. Y., Orr H. T. (2000) Annu Rev. Neurosci. 23, 217–247 - PubMed
-
- Penney J. B., Jr., Vonsattel J. P., MacDonald M. E., Gusella J. F., Myers R. H. (1997) Ann. Neurol. 41, 689–692 - PubMed
-
- Snell R. G., MacMillan J. C., Cheadle J. P., Fenton I., Lazarou L. P., Davies P., MacDonald M. E., Gusella J. F., Harper P. S., Shaw D. J. (1993) Nat. Genet. 4, 393–397 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
