

Somatic sex identity is cell autonomous in the chicken
- PMID: 20220842
- PMCID: PMC3925877
- DOI: 10.1038/nature08852
Somatic sex identity is cell autonomous in the chicken
Abstract
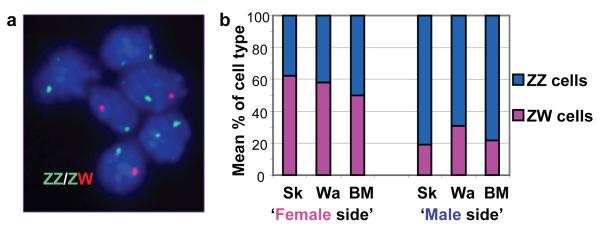
In the mammalian model of sex determination, embryos are considered to be sexually indifferent until the transient action of a sex-determining gene initiates gonadal differentiation. Although this model is thought to apply to all vertebrates, this has yet to be established. Here we have examined three lateral gynandromorph chickens (a rare, naturally occurring phenomenon in which one side of the animal appears male and the other female) to investigate the sex-determining mechanism in birds. These studies demonstrated that gynandromorph birds are genuine male:female chimaeras, and indicated that male and female avian somatic cells may have an inherent sex identity. To test this hypothesis, we transplanted presumptive mesoderm between embryos of reciprocal sexes to generate embryos containing male:female chimaeric gonads. In contrast to the outcome for mammalian mixed-sex chimaeras, in chicken mixed-sex chimaeras the donor cells were excluded from the functional structures of the host gonad. In an example where female tissue was transplanted into a male host, donor cells contributing to the developing testis retained a female identity and expressed a marker of female function. Our study demonstrates that avian somatic cells possess an inherent sex identity and that, in birds, sexual differentiation is substantively cell autonomous.
Figures




Comment in
-
Sex determination: An avian sexual revolution.Nature. 2010 Mar 11;464(7286):171-2. doi: 10.1038/464171a. Nature. 2010. PMID: 20220830 No abstract available.
References
-
- Jost A. Hormonal factors in the sex differentiation of the mammalian foetus. Phil. Trans. R. Soc. Lond. B. 1970;259:119–130. - PubMed
-
- Arnold AP. Sexual differentiation of the zebra finch song system: positive evidence, negative evidence, null hypothesis, and a paradigm shift. J. Neurobiol. 1997;33:572–584. - PubMed
Methods References
-
- Stern CD. Immunocytochemistry of embryonic material. In: Stern CD, Holland PWH, editors. Essential Developmental Biology: A Practical Approach. IRL Press; Oxford, UK: 1993. pp. 193–212.
-
- McQueen HA, et al. CpG islands of chicken are concentrated on microchromosomes. Nature Genetics. 1996;12:321–324. - PubMed
-
- Clinton M, Haines L, Belloir B, Mcbride D. Sexing chick embryos: a rapid and simple protocol. Br. Poultry Sci. 2001;42:134–138. - PubMed
-
- Henrique D, et al. Expression of a Delta homologue in prospective neurons in the chick. Nature. 1995;375:787–790. - PubMed
-
- Clinton M, Miele G, Nandi S, McBride D. Identification of disease markers by differential display: prion disease. In: Liang P, Meade JD, Pardee AB, editors. Differential Display Methods and Protocols. 2nd edition Humana Press; Totowa, NJ: 2006. pp. 157–178. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BBE0154251/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/H012486/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E011276/2/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E015425/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E015425/2/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
