

Kaposi's sarcoma-associated herpesvirus ORF57 protein binds and protects a nuclear noncoding RNA from cellular RNA decay pathways
- PMID: 20221435
- PMCID: PMC2832700
- DOI: 10.1371/journal.ppat.1000799
Kaposi's sarcoma-associated herpesvirus ORF57 protein binds and protects a nuclear noncoding RNA from cellular RNA decay pathways
Abstract
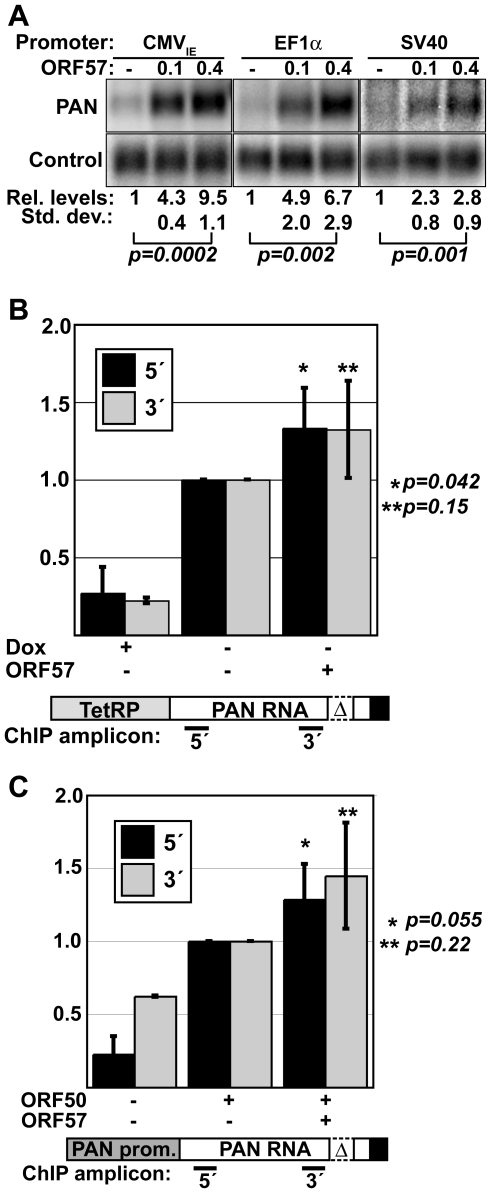
The control of RNA stability is a key determinant in cellular gene expression. The stability of any transcript is modulated through the activity of cis- or trans-acting regulatory factors as well as cellular quality control systems that ensure the integrity of a transcript. As a result, invading viral pathogens must be able to subvert cellular RNA decay pathways capable of destroying viral transcripts. Here we report that the Kaposi's sarcoma-associated herpesvirus (KSHV) ORF57 protein binds to a unique KSHV polyadenylated nuclear RNA, called PAN RNA, and protects it from degradation by cellular factors. ORF57 increases PAN RNA levels and its effects are greatest on unstable alleles of PAN RNA. Kinetic analysis of transcription pulse assays shows that ORF57 protects PAN RNA from a rapid cellular RNA decay process, but ORF57 has little effect on transcription or PAN RNA localization based on chromatin immunoprecipitation and in situ hybridization experiments, respectively. Using a UV cross-linking technique, we further demonstrate that ORF57 binds PAN RNA directly in living cells and we show that binding correlates with function. In addition, we define an ORF57-responsive element (ORE) that is necessary for ORF57 binding to PAN RNA and sufficient to confer ORF57-response to a heterologous intronless beta-globin mRNA, but not its spliced counterparts. We conclude that ORF57 binds to viral transcripts in the nucleus and protects them from a cellular RNA decay pathway. We propose that KSHV ORF57 protein functions to enhance the nuclear stability of intronless viral transcripts by protecting them from a cellular RNA quality control pathway.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







Similar articles
-
Kaposi's sarcoma-associated herpesvirus ORF57 protein protects viral transcripts from specific nuclear RNA decay pathways by preventing hMTR4 recruitment.PLoS Pathog. 2019 Feb 20;15(2):e1007596. doi: 10.1371/journal.ppat.1007596. eCollection 2019 Feb. PLoS Pathog. 2019. PMID: 30785952 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus ORF57 protein enhances mRNA accumulation independently of effects on nuclear RNA export.J Virol. 2007 Sep;81(18):9990-8. doi: 10.1128/JVI.00896-07. Epub 2007 Jul 3. J Virol. 2007. PMID: 17609285 Free PMC article.
-
New insights into the expression and functions of the Kaposi's sarcoma-associated herpesvirus long noncoding PAN RNA.Virus Res. 2016 Jan 2;212:53-63. doi: 10.1016/j.virusres.2015.06.012. Epub 2015 Jun 21. Virus Res. 2016. PMID: 26103097 Free PMC article. Review.
-
Recruitment of the complete hTREX complex is required for Kaposi's sarcoma-associated herpesvirus intronless mRNA nuclear export and virus replication.PLoS Pathog. 2008 Oct;4(10):e1000194. doi: 10.1371/journal.ppat.1000194. Epub 2008 Oct 31. PLoS Pathog. 2008. PMID: 18974867 Free PMC article.
-
Posttranscriptional gene regulation in Kaposi's sarcoma-associated herpesvirus.Adv Appl Microbiol. 2009;68:241-61. doi: 10.1016/S0065-2164(09)01206-4. Adv Appl Microbiol. 2009. PMID: 19426857 Review.
Cited by
-
SAM homeostasis is regulated by CFIm-mediated splicing of MAT2A.Elife. 2021 May 5;10:e64930. doi: 10.7554/eLife.64930. Elife. 2021. PMID: 33949310 Free PMC article.
-
HITS-CLIP analysis uncovers a link between the Kaposi's sarcoma-associated herpesvirus ORF57 protein and host pre-mRNA metabolism.PLoS Pathog. 2015 Feb 24;11(2):e1004652. doi: 10.1371/journal.ppat.1004652. eCollection 2015 Feb. PLoS Pathog. 2015. PMID: 25710169 Free PMC article.
-
Epstein-Barr Virus SM protein utilizes cellular splicing factor SRp20 to mediate alternative splicing.J Virol. 2010 Nov;84(22):11781-9. doi: 10.1128/JVI.01359-10. Epub 2010 Sep 1. J Virol. 2010. PMID: 20810723 Free PMC article.
-
Functional analysis of 3'-UTR hairpins supports a two-tiered model for posttranscriptional regulation of MAT2A by METTL16.RNA. 2023 Nov;29(11):1725-1737. doi: 10.1261/rna.079695.123. Epub 2023 Aug 11. RNA. 2023. PMID: 37567786 Free PMC article.
-
Mutation of a C-terminal motif affects Kaposi's sarcoma-associated herpesvirus ORF57 RNA binding, nuclear trafficking, and multimerization.J Virol. 2011 Aug;85(15):7881-91. doi: 10.1128/JVI.00138-11. Epub 2011 May 18. J Virol. 2011. PMID: 21593148 Free PMC article.
References
-
- Anderson P, Kedersha N. RNA granules: post-transcriptional and epigenetic modulators of gene expression. Nat Rev Mol Cell Biol. 2009;10:430–436. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
