

A non-tree-based comprehensive study of metazoan Hox and ParaHox genes prompts new insights into their origin and evolution
- PMID: 20222951
- PMCID: PMC2842273
- DOI: 10.1186/1471-2148-10-73
A non-tree-based comprehensive study of metazoan Hox and ParaHox genes prompts new insights into their origin and evolution
Abstract
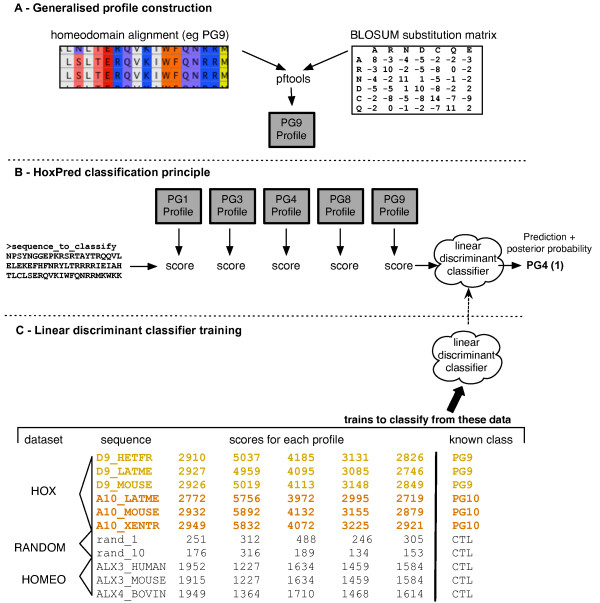
Background: Hox and the closely-related ParaHox genes, which emerged prior to the divergence between cnidarians and bilaterians, are the most well-known members of the ancient genetic toolkit that controls embryonic development across all metazoans. Fundamental questions relative to their origin and evolutionary relationships remain however unresolved. We investigate here the evolution of metazoan Hox and ParaHox genes using the HoxPred program that allows the identification of Hox genes without the need of phylogenetic tree reconstructions.
Results: We show that HoxPred provides an efficient and accurate classification of Hox and ParaHox genes in their respective homology groups, including Hox paralogous groups (PGs). We analyzed more than 10,000 sequences from 310 metazoan species, from 6 genome projects and the complete UniProtKB database. The HoxPred program and all results arranged in the Datab'Hox database are freely available at http://cege.vub.ac.be/hoxpred/. Results for the genome-scale studies are coherent with previous studies, and also brings knowledge on the Hox repertoire and clusters for newly-sequenced species. The unprecedented scale of this study and the use of a non-tree-based approach allows unresolved key questions about Hox and ParaHox genes evolution to be addressed.
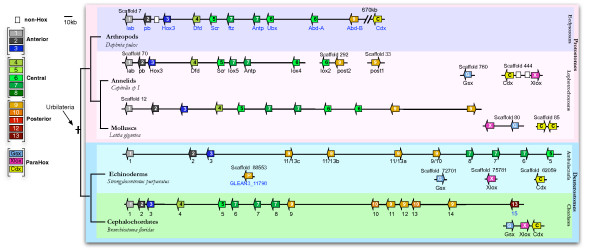
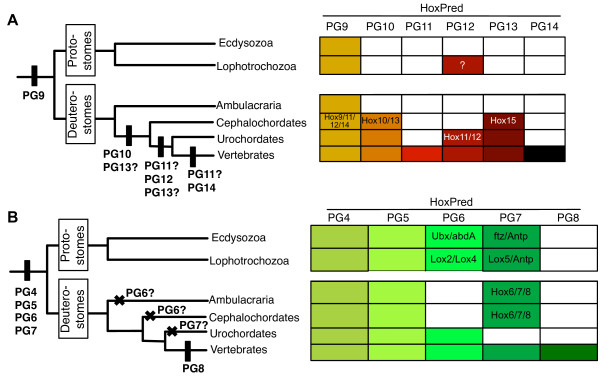
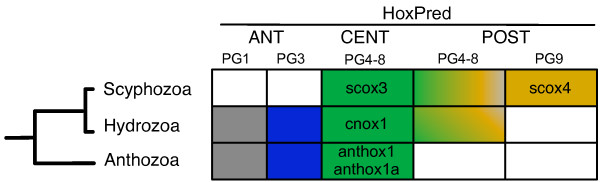
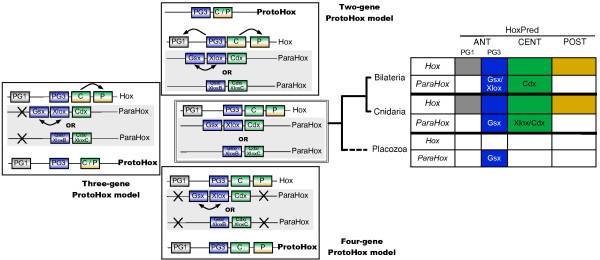
Conclusions: Our analysis suggests that the presence of a single type of Posterior Hox genes (PG9-like) is ancestral to bilaterians, and that new Posterior PGs would have arisen in deuterostomes through independent gene duplications. Four types of Central genes would also be ancestral to bilaterians, with two of them, PG6- and PG7-like that gave rise, in protostomes, to the UbdA- and ftz/Antp/Lox5-type genes, respectively. A fifth type of Central genes (PG8) would have emerged in the vertebrate lineage. Our results also suggest the presence of Anterior (PG1 and PG3), Central and Posterior Hox genes in the cnidarians, supporting an ancestral four-gene Hox cluster. In addition, our data support the relationship of the bilaterian ParaHox genes Gsx and Xlox with PG3, and Cdx with the Central genes. Our study therefore indicates three possible models for the origin of Hox and ParaHox in early metazoans, a two-gene (Anterior/PG3--Central/Posterior), a three-gene (Anterior/PG1, Anterior/PG3 and Central/Posterior), or a four-gene (Anterior/PG1--Anterior/PG3--Central--Posterior) ProtoHox cluster.
Figures
Similar articles
-
More constraint on ParaHox than Hox gene families in early metazoan evolution.Dev Biol. 2009 Apr 15;328(2):173-87. doi: 10.1016/j.ydbio.2009.01.022. Epub 2009 Jan 27. Dev Biol. 2009. PMID: 19389364
-
Minimal ProtoHox cluster inferred from bilaterian and cnidarian Hox complements.Nature. 2006 Aug 10;442(7103):684-7. doi: 10.1038/nature04863. Nature. 2006. PMID: 16900199
-
Do cnidarians have a ParaHox cluster? Analysis of synteny around a Nematostella homeobox gene cluster.Evol Dev. 2008 Nov-Dec;10(6):725-30. doi: 10.1111/j.1525-142X.2008.00286.x. Evol Dev. 2008. PMID: 19021743
-
Hox, ParaHox, ProtoHox: facts and guesses.Heredity (Edinb). 2005 Feb;94(2):145-52. doi: 10.1038/sj.hdy.6800621. Heredity (Edinb). 2005. PMID: 15578045 Review.
-
Evolution of invertebrate deuterostomes and Hox/ParaHox genes.Genomics Proteomics Bioinformatics. 2011 Jun;9(3):77-96. doi: 10.1016/S1672-0229(11)60011-9. Genomics Proteomics Bioinformatics. 2011. PMID: 21802045 Free PMC article. Review.
Cited by
-
The genetic factors of bilaterian evolution.Elife. 2020 Jul 16;9:e45530. doi: 10.7554/eLife.45530. Elife. 2020. PMID: 32672535 Free PMC article.
-
Molecular insights into the origin of the Hox-TALE patterning system.Elife. 2014 Mar 18;3:e01939. doi: 10.7554/eLife.01939. Elife. 2014. PMID: 24642410 Free PMC article.
-
Hox proteins display a common and ancestral ability to diversify their interaction mode with the PBC class cofactors.PLoS Biol. 2012;10(6):e1001351. doi: 10.1371/journal.pbio.1001351. Epub 2012 Jun 26. PLoS Biol. 2012. PMID: 22745600 Free PMC article.
-
Evolution of coding and non-coding genes in HOX clusters of a marsupial.BMC Genomics. 2012 Jun 18;13:251. doi: 10.1186/1471-2164-13-251. BMC Genomics. 2012. PMID: 22708672 Free PMC article.
-
Evolution of Hox gene clusters in deuterostomes.BMC Dev Biol. 2013 Jul 2;13:26. doi: 10.1186/1471-213X-13-26. BMC Dev Biol. 2013. PMID: 23819519 Free PMC article. Review.
References
-
- Burke AC, Nelson CE, Morgan BA, Tabin C. Hox genes and the evolution of vertebrate axial morphology. Development. 1995;121(2):333–346. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
