

Hippocampal replay is not a simple function of experience
- PMID: 20223204
- PMCID: PMC4460981
- DOI: 10.1016/j.neuron.2010.01.034
Hippocampal replay is not a simple function of experience
Abstract
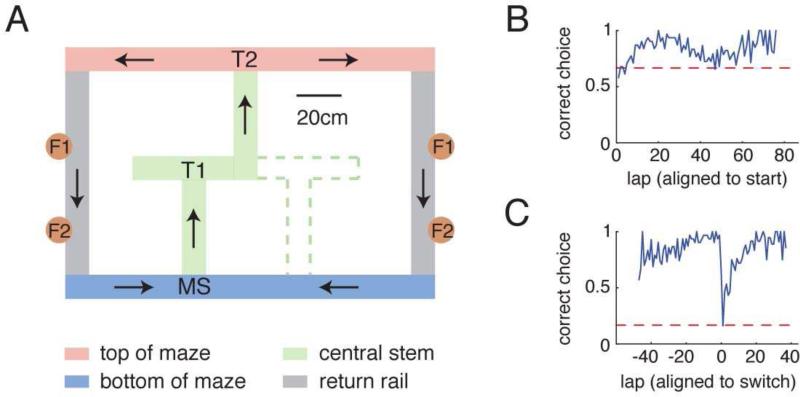
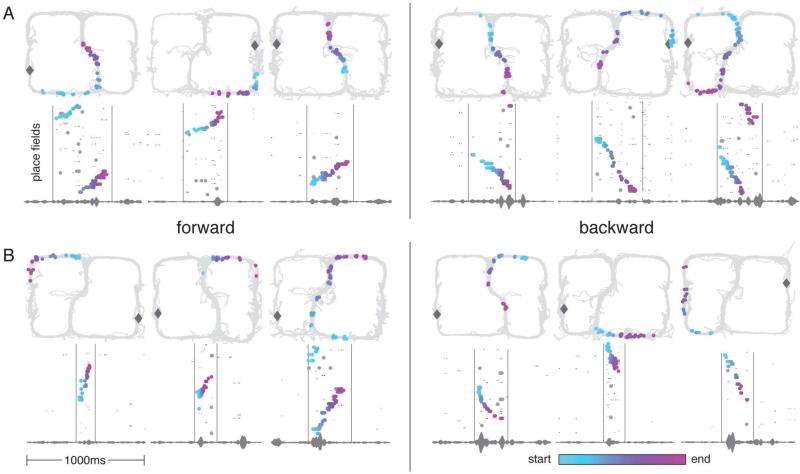
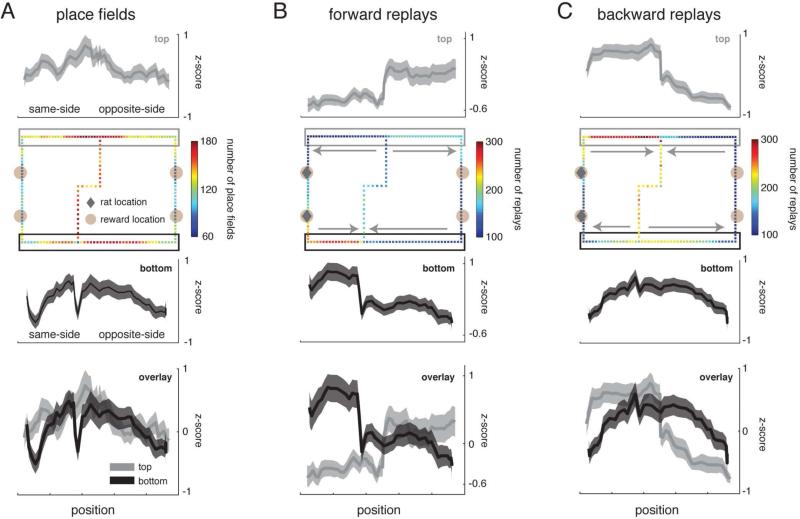
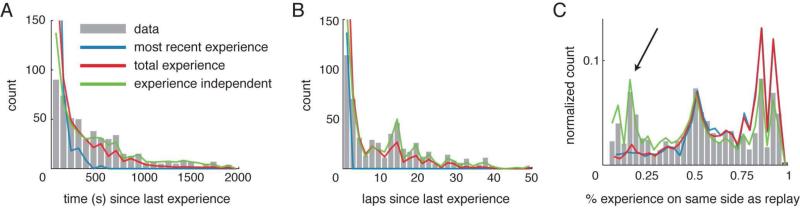
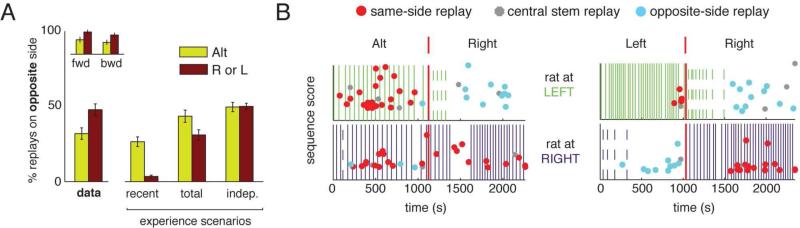
Replay of behavioral sequences in the hippocampus during sharp wave ripple complexes (SWRs) provides a potential mechanism for memory consolidation and the learning of knowledge structures. Current hypotheses imply that replay should straightforwardly reflect recent experience. However, we find these hypotheses to be incompatible with the content of replay on a task with two distinct behavioral sequences (A and B). We observed forward and backward replay of B even when rats had been performing A for >10 min. Furthermore, replay of nonlocal sequence B occurred more often when B was infrequently experienced. Neither forward nor backward sequences preferentially represented highly experienced trajectories within a session. Additionally, we observed the construction of never-experienced novel-path sequences. These observations challenge the idea that sequence activation during SWRs is a simple replay of recent experience. Instead, replay reflected all physically available trajectories within the environment, suggesting a potential role in active learning and maintenance of the cognitive map.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A dual role for hippocampal replay.Neuron. 2010 Mar 11;65(5):582-4. doi: 10.1016/j.neuron.2010.02.022. Neuron. 2010. PMID: 20223195
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
