

Monaural deprivation disrupts development of binaural selectivity in auditory midbrain and cortex
- PMID: 20223206
- PMCID: PMC2849994
- DOI: 10.1016/j.neuron.2010.02.019
Monaural deprivation disrupts development of binaural selectivity in auditory midbrain and cortex
Abstract
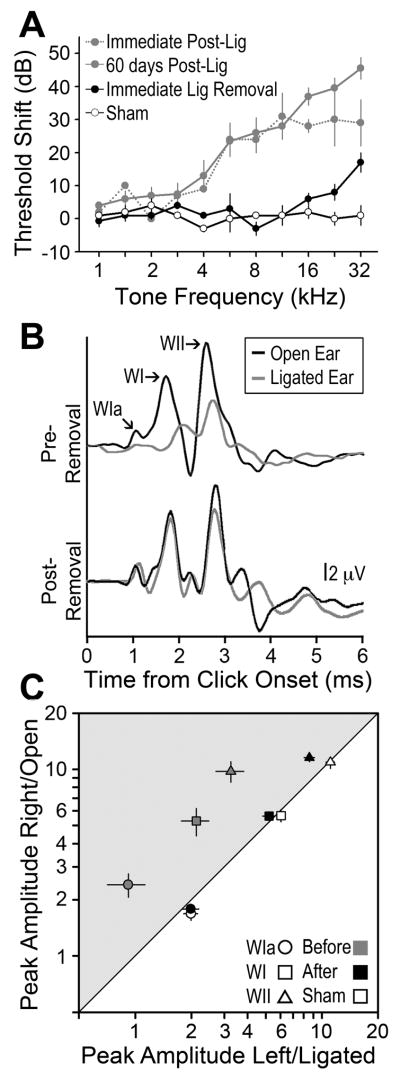
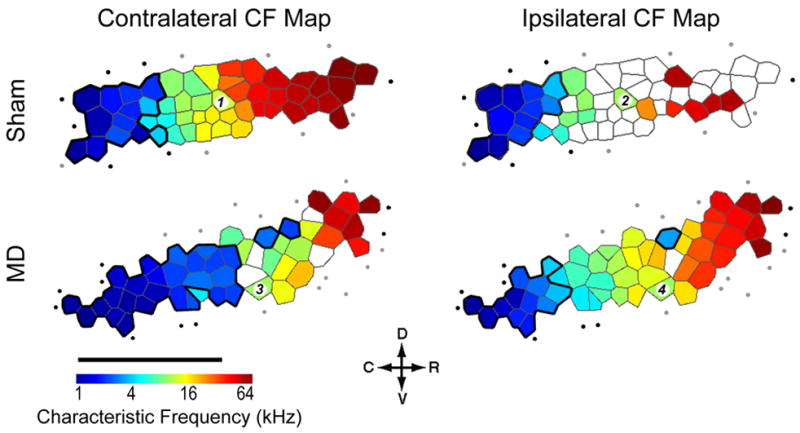
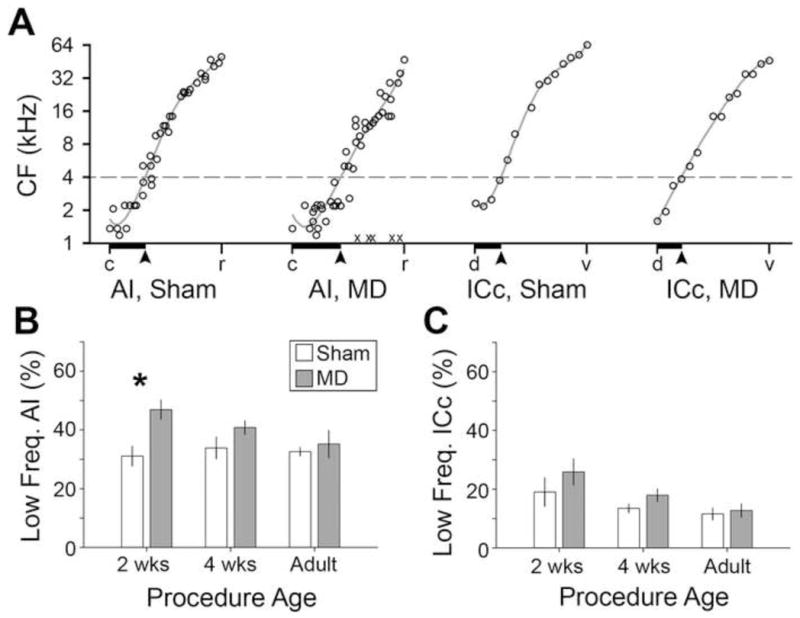
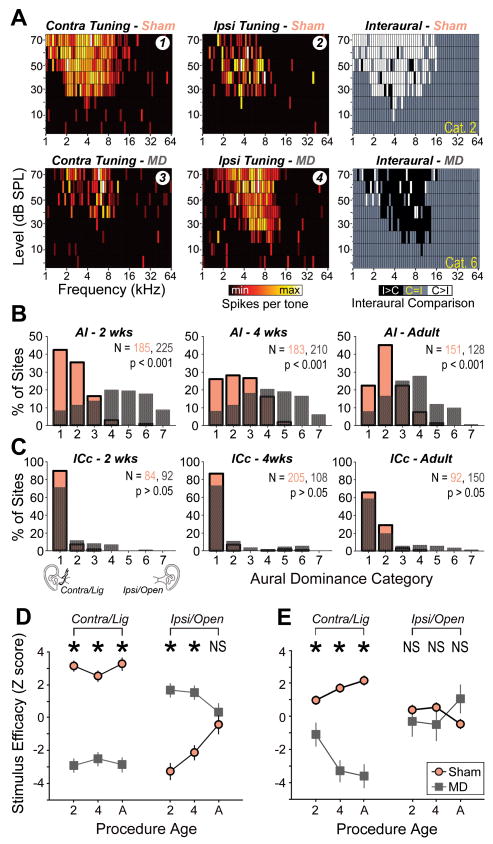
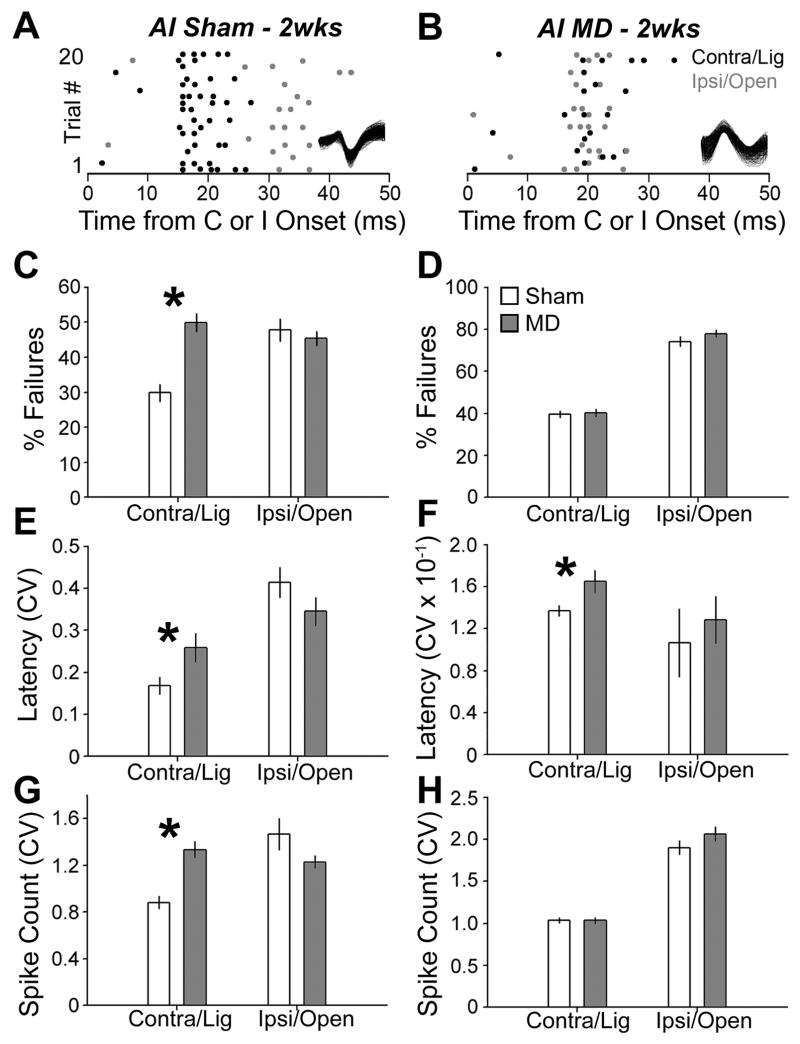
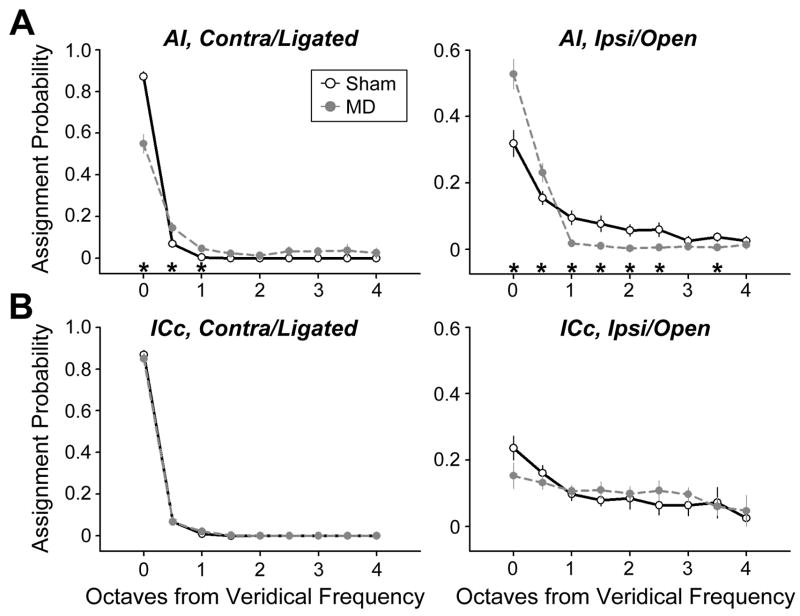
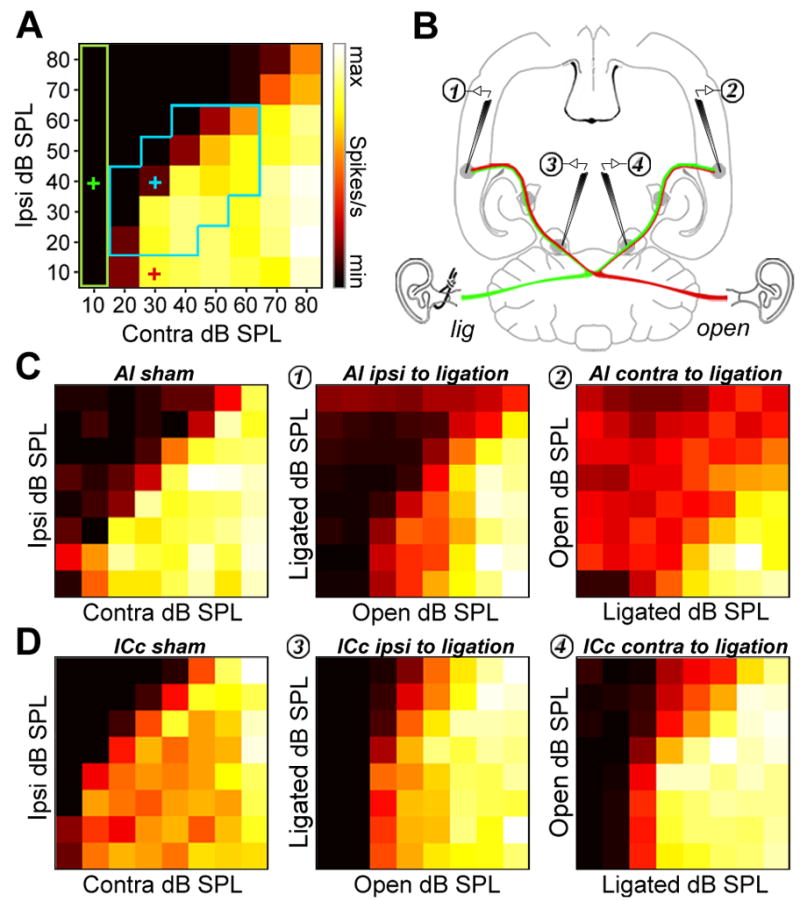
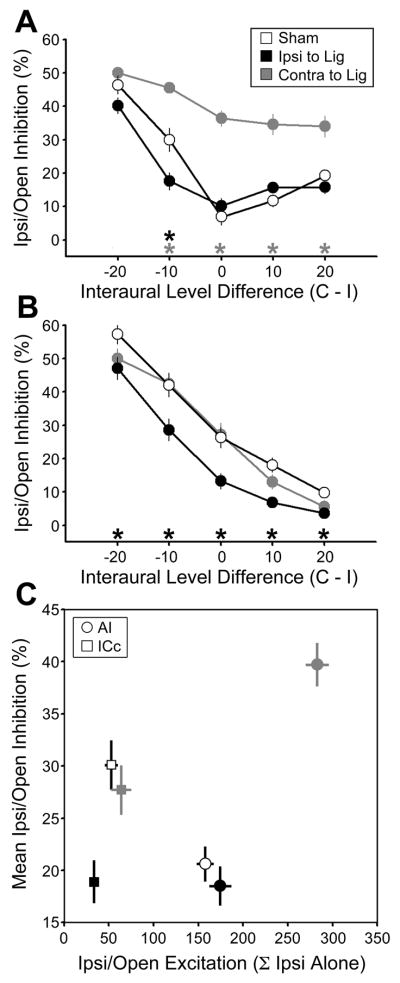
Degraded sensory experience during critical periods of development can have adverse effects on brain function. In the auditory system, conductive hearing loss associated with childhood ear infections can produce long-lasting deficits in auditory perceptual acuity, much like amblyopia in the visual system. Here we explore the neural mechanisms that may underlie "amblyaudio" by inducing reversible monaural deprivation (MD) in infant, juvenile, and adult rats. MD distorted tonotopic maps, weakened the deprived ear's representation, strengthened the open ear's representation, and disrupted binaural integration of interaural level differences (ILD). Bidirectional plasticity effects were strictly governed by critical periods, were more strongly expressed in primary auditory cortex than inferior colliculus, and directly impacted neural coding accuracy. These findings highlight a remarkable degree of competitive plasticity between aural representations and suggest that the enduring perceptual sequelae of childhood hearing loss might be traced to maladaptive plasticity during critical periods of auditory cortex development.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Bendor D, Wang X. Differential neural coding of acoustic flutter within primate auditory cortex. Nat Neurosci. 2007;10:763–771. - PubMed
-
- Chang EF, Merzenich MM. Environmental noise retards auditory cortical development. Science. 2003;300:498–502. - PubMed
-
- Clements M, Kelly JB. Auditory spatial responses of young guinea pigs (Cavia porcellus) during and after ear blocking. J Comp Physiol Psychol. 1978;92:34–44. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
