

The Mi-2-like Smed-CHD4 gene is required for stem cell differentiation in the planarian Schmidtea mediterranea
- PMID: 20223763
- PMCID: PMC2847463
- DOI: 10.1242/dev.042051
The Mi-2-like Smed-CHD4 gene is required for stem cell differentiation in the planarian Schmidtea mediterranea
Abstract
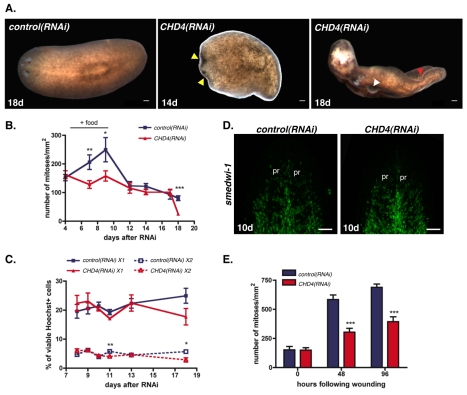
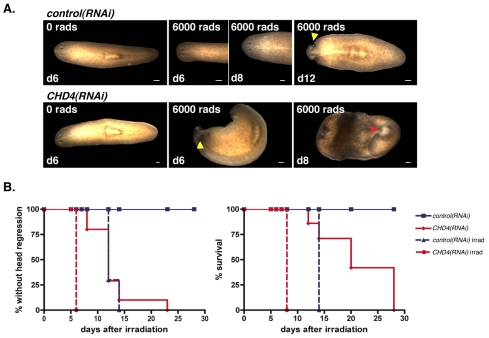
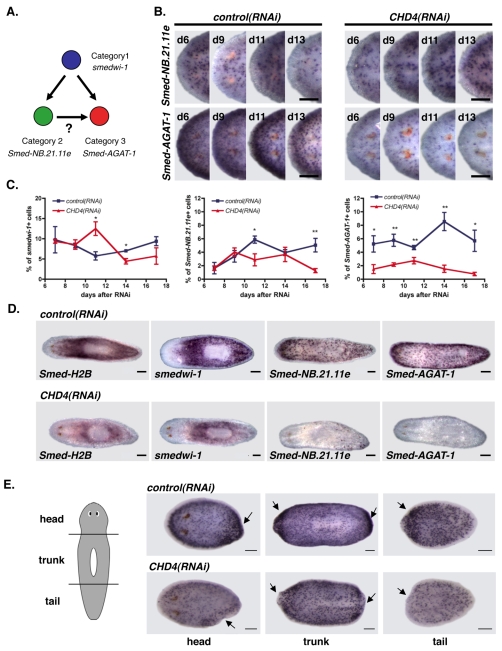
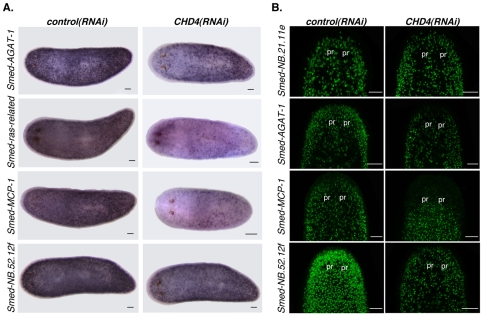
Freshwater planarians are able to regenerate any missing part of their body and have extensive tissue turnover because of the action of dividing cells called neoblasts. Neoblasts provide an excellent system for in vivo study of adult stem cell biology. We identified the Smed-CHD4 gene, which is predicted to encode a chromatin-remodeling protein similar to CHD4/Mi-2 proteins, as required for planarian regeneration and tissue homeostasis. Following inhibition of Smed-CHD4 with RNA interference (RNAi), neoblast numbers were initially normal, despite an inability of the animals to regenerate. However, the proliferative response of neoblasts to amputation or growth stimulation in Smed-CHD4(RNAi) animals was diminished. Smed-CHD4(RNAi) animals displayed a dramatic reduction in the numbers of certain neoblast progeny cells. Smed-CHD4 was required for the formation of these neoblast progeny cells. Together, these results indicate that Smed-CHD4 is required for neoblasts to produce progeny cells committed to differentiation in order to control tissue turnover and regeneration and suggest a crucial role for CHD4 proteins in stem cell differentiation.
Figures


Similar articles
-
A Bruno-like gene is required for stem cell maintenance in planarians.Dev Cell. 2006 Aug;11(2):159-69. doi: 10.1016/j.devcel.2006.06.004. Dev Cell. 2006. PMID: 16890156
-
SMEDWI-2 is a PIWI-like protein that regulates planarian stem cells.Science. 2005 Nov 25;310(5752):1327-30. doi: 10.1126/science.1116110. Science. 2005. PMID: 16311336
-
Smed-egfr-4 is required for planarian eye regeneration.Int J Dev Biol. 2019;63(1-2):9-15. doi: 10.1387/ijdb.180361fc. Int J Dev Biol. 2019. PMID: 30919917
-
Decoding Stem Cells: An Overview on Planarian Stem Cell Heterogeneity and Lineage Progression.Biomolecules. 2021 Oct 17;11(10):1532. doi: 10.3390/biom11101532. Biomolecules. 2021. PMID: 34680165 Free PMC article. Review.
-
Planarians, a tale of stem cells.Cell Mol Life Sci. 2008 Jan;65(1):16-23. doi: 10.1007/s00018-007-7426-y. Cell Mol Life Sci. 2008. PMID: 18030424 Free PMC article. Review.
Cited by
-
In situ hybridization protocol for enhanced detection of gene expression in the planarian Schmidtea mediterranea.BMC Dev Biol. 2013 Mar 12;13:8. doi: 10.1186/1471-213X-13-8. BMC Dev Biol. 2013. PMID: 23497040 Free PMC article.
-
Clonogenic neoblasts are pluripotent adult stem cells that underlie planarian regeneration.Science. 2011 May 13;332(6031):811-6. doi: 10.1126/science.1203983. Science. 2011. PMID: 21566185 Free PMC article.
-
Single-cell analysis reveals functionally distinct classes within the planarian stem cell compartment.Cell Stem Cell. 2014 Sep 4;15(3):326-339. doi: 10.1016/j.stem.2014.06.007. Epub 2014 Jul 10. Cell Stem Cell. 2014. PMID: 25017721 Free PMC article.
-
Stochastic cell-intrinsic stem cell decisions control colony growth in planarians.Elife. 2025 Jul 23;13:RP100885. doi: 10.7554/eLife.100885. Elife. 2025. PMID: 40699888 Free PMC article.
-
Establishment of a new method to isolate viable x-ray-sensitive cells from planarian by fluorescence-activated cell sorting.Dev Growth Differ. 2023 Dec;65(9):577-590. doi: 10.1111/dgd.12886. Epub 2023 Sep 5. Dev Growth Differ. 2023. PMID: 37596847 Free PMC article.
References
-
- Ahringer J. (2000). NuRD and SIN3 histone deacetylase complexes in development. Trends Genet. 16, 351-356 - PubMed
-
- Baguñà J. (1976). Mitosis in the intact and regenerating planarian Dugesia mediterranea n.sp. I. Mitotic studies during growth, feeding and starvation. J. Exp. Zool. 195, 53-64
-
- Bardeen C. R., Baetjer F. H. (1904). The inhibitive action of the Roentgen rays on regeneration in planarians. J. Exp. Zool. 1, 191-195
-
- Becker P. B., Horz W. (2002). ATP-dependent nucleosome remodeling. Annu. Rev. Biochem. 71, 247-273 - PubMed
-
- Bibikova M., Laurent L. C., Ren B., Loring J. F., Fan J. B. (2008). Unraveling epigenetic regulation in embryonic stem cells. Cell Stem Cell 2, 123-134 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
