

Phosphorylation-induced conformational changes in the retinoblastoma protein inhibit E2F transactivation domain binding
- PMID: 20223825
- PMCID: PMC2871496
- DOI: 10.1074/jbc.M110.108167
Phosphorylation-induced conformational changes in the retinoblastoma protein inhibit E2F transactivation domain binding
Abstract
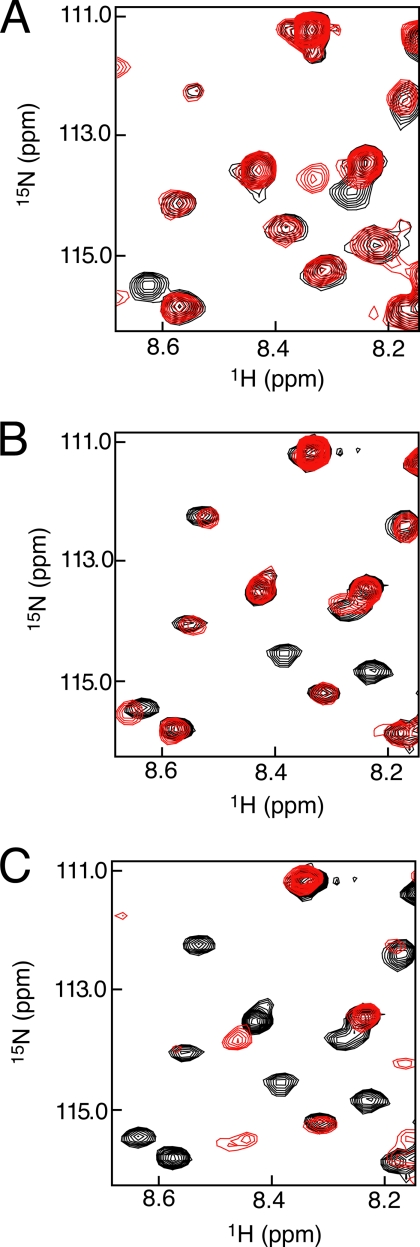
Inactivation of the retinoblastoma protein (Rb) through phosphorylation is an important step in promoting cell cycle progression, and hyperphosphorylated Rb is commonly found in tumors. Rb phosphorylation prevents its association with the E2F transcription factor; however, the molecular basis for complex inhibition has not been established. We identify here the key phosphorylation events and conformational changes that occur in Rb to inhibit the specific association between the E2F transactivation domain (E2F(TD)) and the Rb pocket domain. Calorimetry assays demonstrate that phosphorylation of Rb reduces the affinity of E2F(TD) binding approximately 250-fold and that phosphorylation at Ser(608)/Ser(612) and Thr(356)/Thr(373) is necessary and sufficient for this effect. An NMR assay identifies phosphorylation-driven conformational changes in Rb that directly inhibit E2F(TD) binding. We find that phosphorylation at Ser(608)/Ser(612) promotes an intramolecular association between a conserved sequence in the flexible pocket linker and the pocket domain of Rb that occludes the E2F(TD) binding site. We also find that phosphorylation of Thr(356)/Thr(373) inhibits E2F(TD) binding in a manner that requires the Rb N-terminal domain. Taken together, our results suggest two distinct mechanisms for how phosphorylation of Rb modulates association between E2F(TD) and the Rb pocket and describe for the first time a function for the structured N-terminal domain in Rb inactivation.
Figures




References
-
- Sherr C. J. (1996) Science 274, 1672–1677 - PubMed
-
- Weinberg R. A. (1995) Cell 81, 323–330 - PubMed
-
- Adams P. D. (2001) Biochim. Biophys. Acta 1471, M123–133 - PubMed
-
- Buchkovich K., Duffy L. A., Harlow E. (1989) Cell 58, 1097–1105 - PubMed
-
- Chen P. L., Scully P., Shew J. Y., Wang J. Y., Lee W. H. (1989) Cell 58, 1193–1198 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
