

Restriction of receptor movement alters cellular response: physical force sensing by EphA2
- PMID: 20223987
- PMCID: PMC2895569
- DOI: 10.1126/science.1181729
Restriction of receptor movement alters cellular response: physical force sensing by EphA2
Abstract
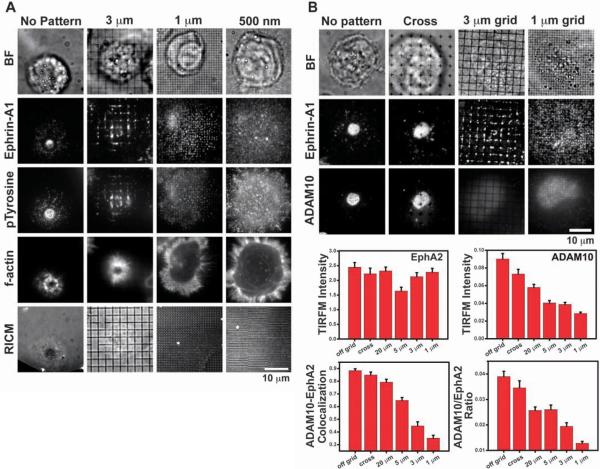
Activation of the EphA2 receptor tyrosine kinase by ephrin-A1 ligands presented on apposed cell surfaces plays important roles in development and exhibits poorly understood functional alterations in cancer. We reconstituted this intermembrane signaling geometry between live EphA2-expressing human breast cancer cells and supported membranes displaying laterally mobile ephrin-A1. Receptor-ligand binding, clustering, and subsequent lateral transport within this junction were observed. EphA2 transport can be blocked by physical barriers nanofabricated onto the underlying substrate. This physical reorganization of EphA2 alters the cellular response to ephrin-A1, as observed by changes in cytoskeleton morphology and recruitment of a disintegrin and metalloprotease 10. Quantitative analysis of receptor-ligand spatial organization across a library of 26 mammary epithelial cell lines reveals characteristic differences that strongly correlate with invasion potential. These observations reveal a mechanism for spatio-mechanical regulation of EphA2 signaling pathways.
Figures



Comment in
-
Biophysics. Enforcing order on signaling.Science. 2010 Mar 12;327(5971):1335-6. doi: 10.1126/science.1187865. Science. 2010. PMID: 20223974 No abstract available.
-
Mechanotransduction: using the force.Nat Rev Mol Cell Biol. 2010 May;11(5):311. doi: 10.1038/nrm2884. Nat Rev Mol Cell Biol. 2010. PMID: 20419888
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
