

A divalent FHA/BRCT-binding mechanism couples the MRE11-RAD50-NBS1 complex to damaged chromatin
- PMID: 20224574
- PMCID: PMC2868538
- DOI: 10.1038/embor.2010.30
A divalent FHA/BRCT-binding mechanism couples the MRE11-RAD50-NBS1 complex to damaged chromatin
Abstract
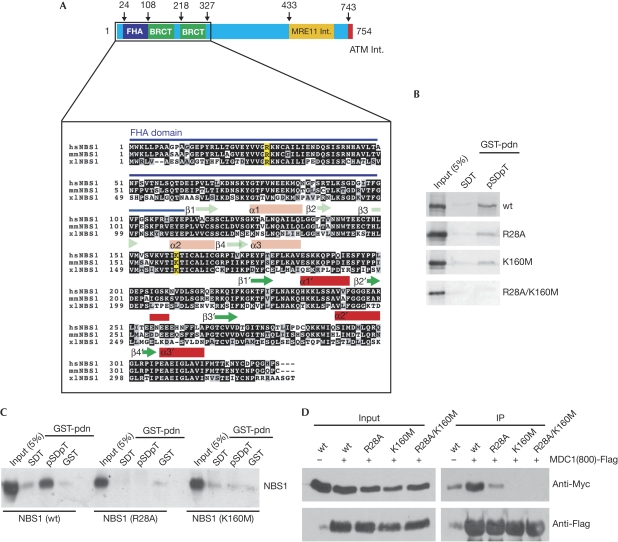
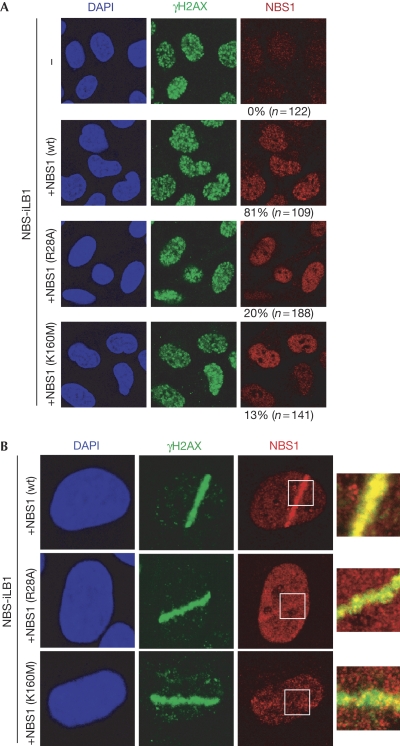
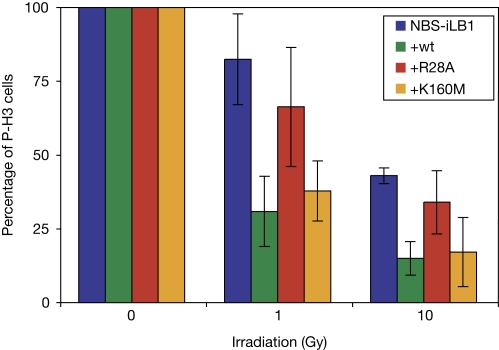
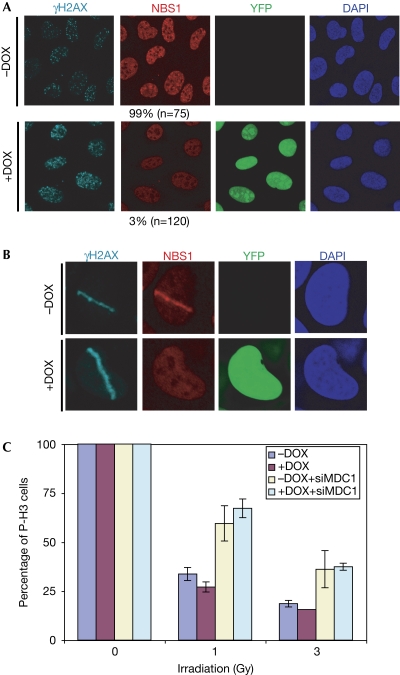
The MRE11-RAD50-NBS1 (MRN) complex accumulates at sites of DNA double-strand breaks in large chromatin domains flanking the lesion site. The mechanism of MRN accumulation involves direct binding of the Nijmegen breakage syndrome 1 (NBS1) subunit to phosphorylated mediator of the DNA damage checkpoint 1 (MDC1), a large nuclear adaptor protein that interacts directly with phosphorylated H2AX. NBS1 contains an FHA domain and two BRCT domains at its amino terminus. Here, we show that both of these domains participate in the interaction with phosphorylated MDC1. Point mutations in key amino acid residues of either the FHA or the BRCT domains compromise the interaction with MDC1 and lead to defects in MRN accumulation at sites of DNA damage. Surprisingly, only mutation in the FHA domain, but not in the BRCT domains, yields a G2/M checkpoint defect, indicating that MDC1-dependent chromatin accumulation of the MRN complex at sites of DNA breaks is not required for G2/M checkpoint activation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Phospho-dependent interactions between NBS1 and MDC1 mediate chromatin retention of the MRN complex at sites of DNA damage.EMBO Rep. 2008 Aug;9(8):795-801. doi: 10.1038/embor.2008.103. Epub 2008 Jun 27. EMBO Rep. 2008. PMID: 18583988 Free PMC article.
-
Constitutive phosphorylation of MDC1 physically links the MRE11-RAD50-NBS1 complex to damaged chromatin.J Cell Biol. 2008 Apr 21;181(2):227-40. doi: 10.1083/jcb.200709008. Epub 2008 Apr 14. J Cell Biol. 2008. PMID: 18411308 Free PMC article.
-
MDC1 regulates intra-S-phase checkpoint by targeting NBS1 to DNA double-strand breaks.Proc Natl Acad Sci U S A. 2008 Aug 12;105(32):11200-5. doi: 10.1073/pnas.0802885105. Epub 2008 Aug 4. Proc Natl Acad Sci U S A. 2008. PMID: 18678890 Free PMC article.
-
Mre11-Rad50-Nbs1 is a keystone complex connecting DNA repair machinery, double-strand break signaling, and the chromatin template.Biochem Cell Biol. 2007 Aug;85(4):509-20. doi: 10.1139/O07-069. Biochem Cell Biol. 2007. PMID: 17713585 Review.
-
Molecular mechanism of the recruitment of NBS1/hMRE11/hRAD50 complex to DNA double-strand breaks: NBS1 binds to gamma-H2AX through FHA/BRCT domain.J Radiat Res. 2004 Dec;45(4):473-8. doi: 10.1269/jrr.45.473. J Radiat Res. 2004. PMID: 15635255 Review.
Cited by
-
DNA folds threaten genetic stability and can be leveraged for chemotherapy.RSC Chem Biol. 2020 Sep 30;2(1):47-76. doi: 10.1039/d0cb00151a. eCollection 2021 Feb 1. RSC Chem Biol. 2020. PMID: 35340894 Free PMC article. Review.
-
The molecular details of a novel phosphorylation-dependent interaction between MRN and the SOSS complex.Protein Sci. 2023 Oct;32(10):e4782. doi: 10.1002/pro.4782. Protein Sci. 2023. PMID: 37705456 Free PMC article.
-
Structural studies of DNA end detection and resection in homologous recombination.Cold Spring Harb Perspect Biol. 2014 Jul 31;6(10):a017962. doi: 10.1101/cshperspect.a017962. Cold Spring Harb Perspect Biol. 2014. PMID: 25081516 Free PMC article. Review.
-
Envisioning the dynamics and flexibility of Mre11-Rad50-Nbs1 complex to decipher its roles in DNA replication and repair.Prog Biophys Mol Biol. 2015 Mar;117(2-3):182-193. doi: 10.1016/j.pbiomolbio.2014.12.004. Epub 2015 Jan 7. Prog Biophys Mol Biol. 2015. PMID: 25576492 Free PMC article. Review.
-
Identification of the interactors of human nibrin (NBN) and of its 26 kDa and 70 kDa fragments arising from the NBN 657del5 founder mutation.PLoS One. 2014 Dec 8;9(12):e114651. doi: 10.1371/journal.pone.0114651. eCollection 2014. PLoS One. 2014. PMID: 25485873 Free PMC article.
References
-
- Becker E, Meyer V, Madaoui H, Guerois R (2006) Detection of a tandem BRCT in Nbs1 and Xrs2 with functional implications in the DNA damage response. Bioinformatics 22: 1289–1292 - PubMed
-
- Cerosaletti KM, Concannon P (2003) Nibrin forkhead-associated domain and breast cancer C-terminal domain are both required for nuclear focus formation and phosphorylation. J Biol Chem 278: 21944–21951 - PubMed
-
- D'Amours D, Jackson SP (2002) The Mre11 complex: at the crossroads of DNA repair and checkpoint signalling. Nat Rev Mol Cell Biol 3: 317–327 - PubMed
-
- Difilippantonio S, Nussenzweig A (2007) The NBS1-ATM connection revisited. Cell Cycle 6: 2366–2370 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
