

GCN5-dependent acetylation of HIV-1 integrase enhances viral integration
- PMID: 20226045
- PMCID: PMC2848186
- DOI: 10.1186/1742-4690-7-18
GCN5-dependent acetylation of HIV-1 integrase enhances viral integration
Abstract
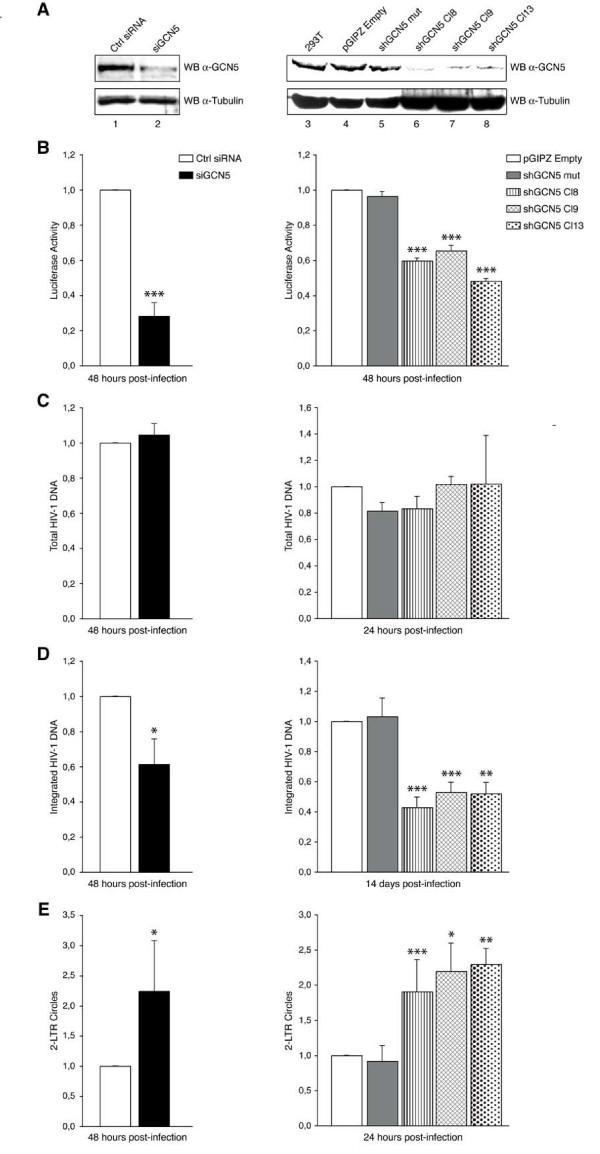
Background: An essential event during the replication cycle of HIV-1 is the integration of the reverse transcribed viral DNA into the host cellular genome. Our former report revealed that HIV-1 integrase (IN), the enzyme that catalyzes the integration reaction, is positively regulated by acetylation mediated by the histone acetyltransferase (HAT) p300.
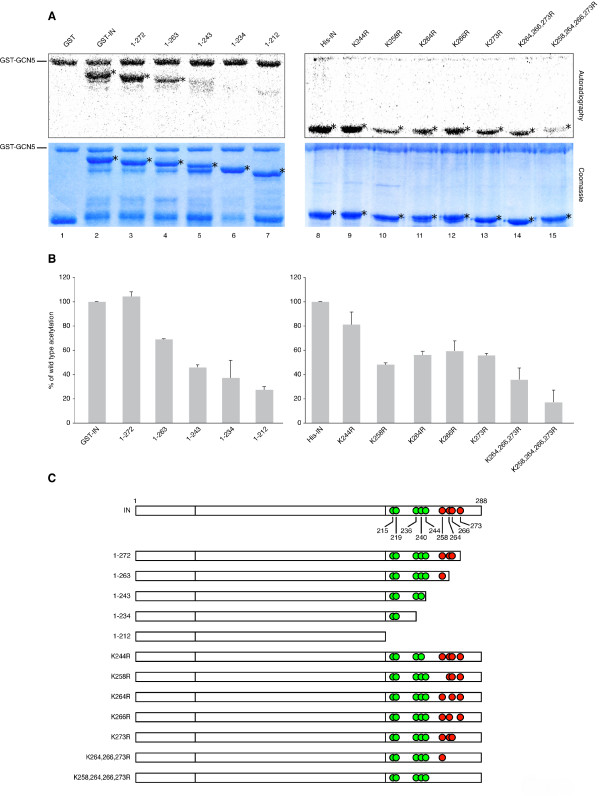
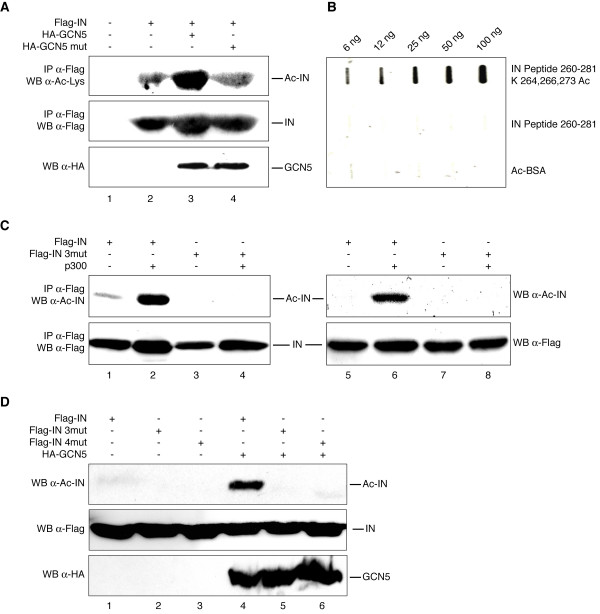
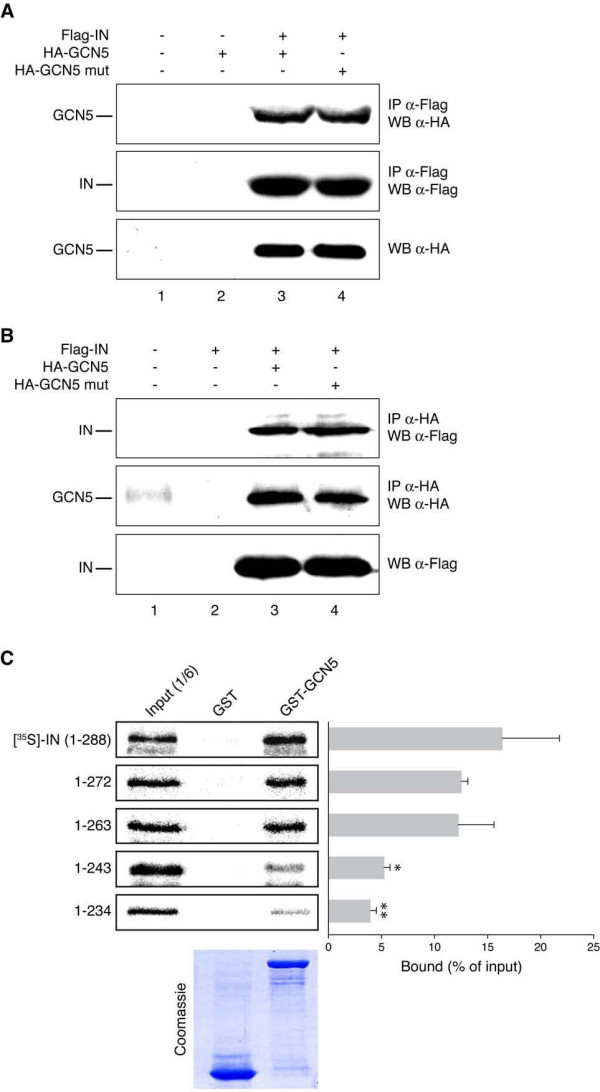
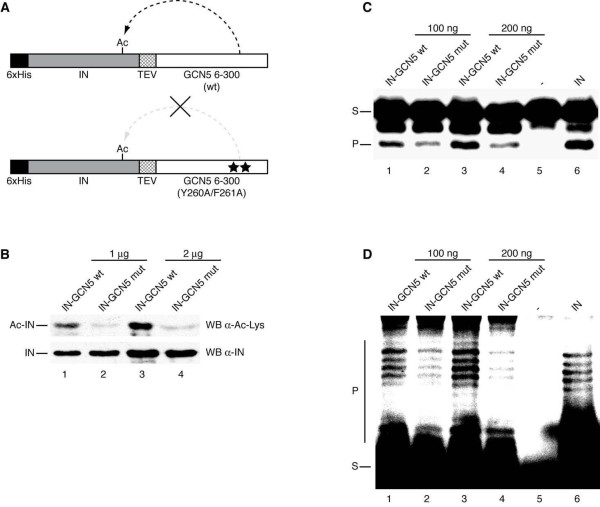
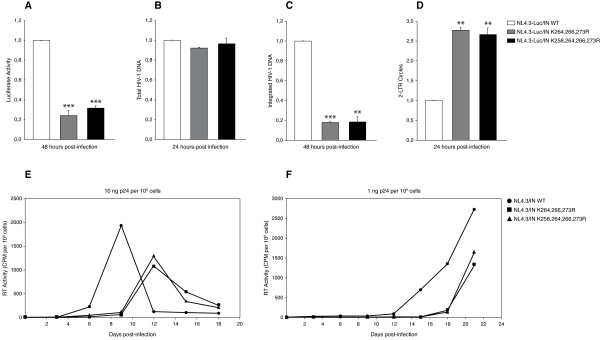
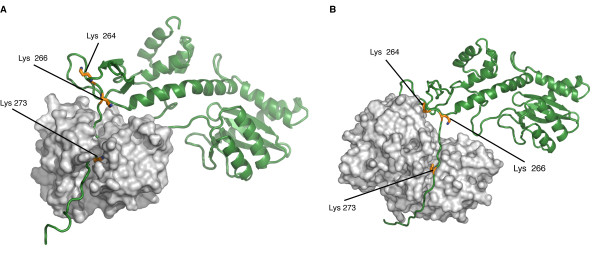
Results: In this study we demonstrate that another cellular HAT, GCN5, acetylates IN leading to enhanced 3'-end processing and strand transfer activities. GCN5 participates in the integration step of HIV-1 replication cycle as demonstrated by the reduced infectivity, due to inefficient provirus formation, in GCN5 knockdown cells. Within the C-terminal domain of IN, four lysines (K258, K264, K266, and K273) are targeted by GCN5 acetylation, three of which (K264, K266, and K273) are also modified by p300. Replication analysis of HIV-1 clones carrying substitutions at the IN lysines acetylated by both GCN5 and p300, or exclusively by GCN5, demonstrated that these residues are required for efficient viral integration. In addition, a comparative analysis of the replication efficiencies of the IN triple- and quadruple-mutant viruses revealed that even though the lysines targeted by both GCN5 and p300 are required for efficient virus integration, the residue exclusively modified by GCN5 (K258) does not affect this process.
Conclusions: The results presented here further demonstrate the relevance of IN post-translational modification by acetylation, which results from the catalytic activities of multiple HATs during the viral replication cycle. Finally, this study contributes to clarifying the recent debate raised on the role of IN acetylated lysines during HIV-1 infection.
Figures
Similar articles
-
Acetylation of HIV-1 integrase by p300 regulates viral integration.EMBO J. 2005 Sep 7;24(17):3070-81. doi: 10.1038/sj.emboj.7600770. Epub 2005 Aug 11. EMBO J. 2005. PMID: 16096645 Free PMC article.
-
Identification of cellular factors binding to acetylated HIV-1 integrase.Amino Acids. 2011 Nov;41(5):1137-45. doi: 10.1007/s00726-009-0444-3. Epub 2009 Dec 18. Amino Acids. 2011. PMID: 20016921
-
Lysine acetylation sites in bovine foamy virus transactivator BTas are important for its DNA binding activity.Virology. 2011 Sep 15;418(1):21-6. doi: 10.1016/j.virol.2011.07.003. Epub 2011 Aug 3. Virology. 2011. PMID: 21813148
-
HIV-1 integration: an interplay between HIV-1 integrase, cellular and viral proteins.AIDS Rev. 2005 Jan-Mar;7(1):26-43. AIDS Rev. 2005. PMID: 15875659 Review.
-
Molecular mechanisms of HIV integration and therapeutic intervention.Expert Rev Mol Med. 2007 Feb 26;9(6):1-19. doi: 10.1017/S1462399407000257. Expert Rev Mol Med. 2007. PMID: 17320002 Review.
Cited by
-
Complex Relationships between HIV-1 Integrase and Its Cellular Partners.Int J Mol Sci. 2022 Oct 15;23(20):12341. doi: 10.3390/ijms232012341. Int J Mol Sci. 2022. PMID: 36293197 Free PMC article. Review.
-
The human antiviral factor TRIM11 is under the regulation of HIV-1 Vpr.PLoS One. 2014 Aug 8;9(8):e104269. doi: 10.1371/journal.pone.0104269. eCollection 2014. PLoS One. 2014. PMID: 25105968 Free PMC article.
-
Protein Acetylation Going Viral: Implications in Antiviral Immunity and Viral Infection.Int J Mol Sci. 2022 Sep 25;23(19):11308. doi: 10.3390/ijms231911308. Int J Mol Sci. 2022. PMID: 36232610 Free PMC article. Review.
-
Involvement of Human Cellular Proteins and Structures in Realization of the HIV Life Cycle: A Comprehensive Review, 2024.Viruses. 2024 Oct 29;16(11):1682. doi: 10.3390/v16111682. Viruses. 2024. PMID: 39599797 Free PMC article. Review.
-
Role of host-encoded proteins in restriction of retroviral integration.Front Microbiol. 2012 Jun 22;3:227. doi: 10.3389/fmicb.2012.00227. eCollection 2012. Front Microbiol. 2012. PMID: 22737148 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
