

In vivo generation of beta-cell-like cells from CD34(+) cells differentiated from human embryonic stem cells
- PMID: 20227460
- PMCID: PMC2880573
- DOI: 10.1016/j.exphem.2010.03.002
In vivo generation of beta-cell-like cells from CD34(+) cells differentiated from human embryonic stem cells
Abstract
Objective: CD34(+) cells, present within the bone marrow, have previously been shown to possess pancreatic endocrine potential. Based on this observation, we explored the capacity of CD34(+) cells derived in culture from the differentiation of human embryonic stem cells (hESC), for their in vivo pancreatic endocrine capacity.
Materials and methods: Sheep were transplanted with hESC-derived CD34(+) cells, as well as nonsorted differentiated cultures. Transplantations were carried out with in utero intraperitoneal injections prior to development of the immune system in the fetus so that tolerance toward foreign antigens was acquired during gestation and persisted in the adult.
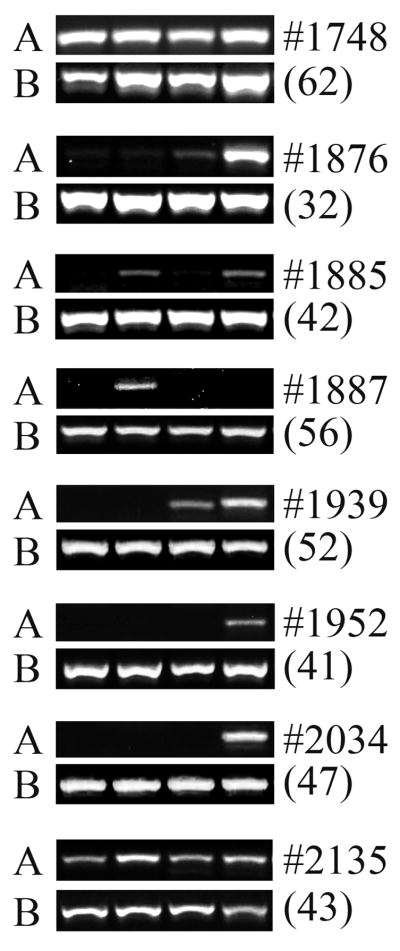
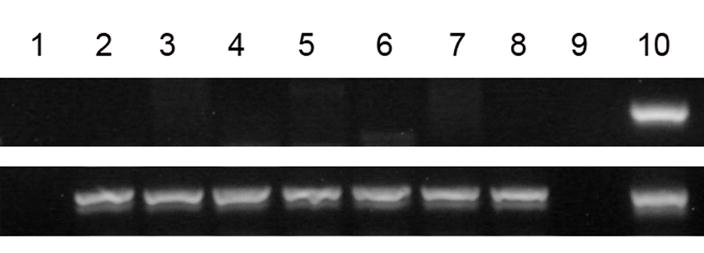
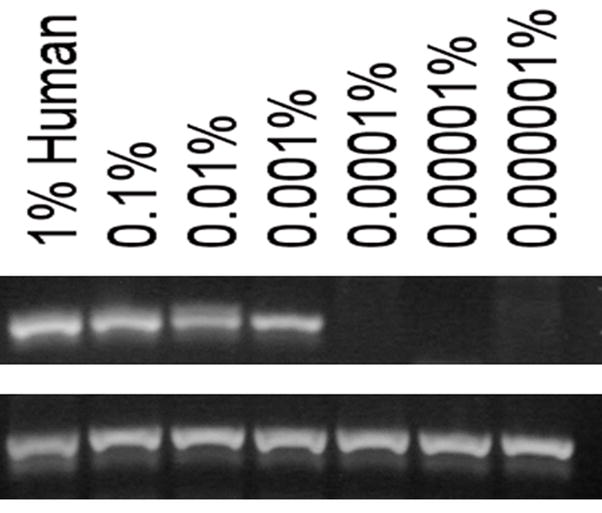
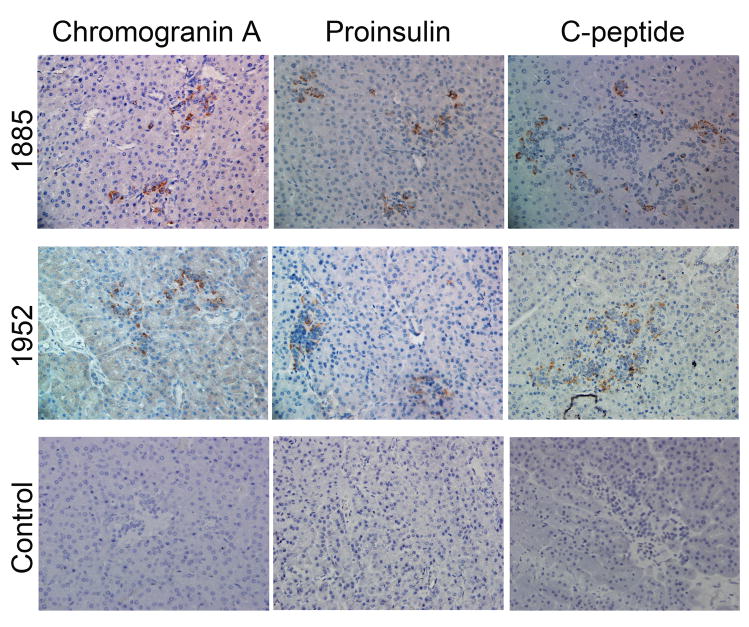
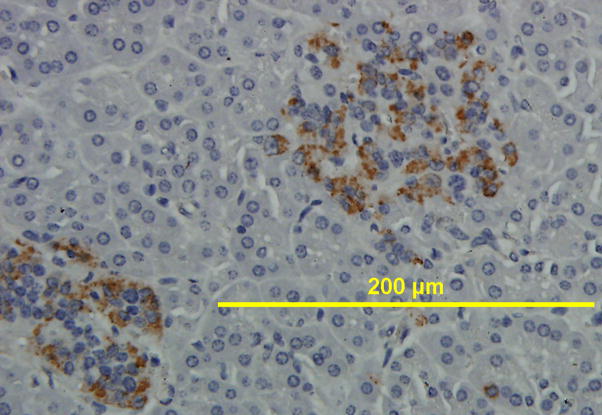
Results: All cell populations that were tested demonstrated human cellular activity and long-term presence up to 5 years. However, the in vivo beta-cell-like activity achieved from the transplantation of the sorted CD34(+) cell population was not augmented by transplanting the entire cell population from which the CD34(+) cells were isolated. Human DNA and insulin messenger RNA were detected in sheep pancreases. An average of 1.51 ng/mL human C-peptide was detected in serum from eight animals transplanted with differentiated cell populations and assayed up to 55 months posttransplantation. Transplantation of as few as 23,500 cells resulted in long-term sustainable beta-cell-like activity. Teratomas were absent in the transplanted animals.
Conclusion: Our data suggest that hESC-derived CD34(+) cells have a potential for long-term in vivo endocrine cellular activity that could prove useful in regenerative medicine. Because the same cell population has previously been shown to contain hematopoietic potential, it could be used for the induction of immunological tolerance and bone marrow chimerism prior to cellular therapy for diabetes.
Copyright 2010 ISEH - Society for Hematology and Stem Cells. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
No financial interest/relationships with financial interest relating to the topic of this article have been declared.
Figures


Similar articles
-
Generation of CD34+ cells from human embryonic stem cells using a clinically applicable methodology and engraftment in the fetal sheep model.Exp Hematol. 2013 Aug;41(8):749-758.e5. doi: 10.1016/j.exphem.2013.04.003. Epub 2013 Apr 20. Exp Hematol. 2013. PMID: 23612043 Free PMC article.
-
In vivo characterization of transplanted human embryonic stem cell-derived pancreatic endocrine islet cells.Cell Prolif. 2008 Dec;41(6):843-858. doi: 10.1111/j.1365-2184.2008.00564.x. Cell Prolif. 2008. PMID: 19040565 Free PMC article.
-
Persistence of human multilineage, self-renewing lymphohematopoietic stem cells in chimeric sheep.Blood. 1993 Dec 1;82(11):3333-42. Blood. 1993. PMID: 7694681
-
Directed differentiation of human embryonic stem cells into the pancreatic endocrine lineage.Stem Cells Dev. 2007 Aug;16(4):561-78. doi: 10.1089/scd.2007.0029. Stem Cells Dev. 2007. PMID: 17784830
-
Bioluminescent imaging demonstrates that transplanted human embryonic stem cell-derived CD34(+) cells preferentially develop into endothelial cells.Stem Cells. 2009 Nov;27(11):2675-85. doi: 10.1002/stem.204. Stem Cells. 2009. PMID: 19711457 Free PMC article.
Cited by
-
Recent Advances in Disease Modeling and Drug Discovery for Diabetes Mellitus Using Induced Pluripotent Stem Cells.Int J Mol Sci. 2016 Feb 19;17(2):256. doi: 10.3390/ijms17020256. Int J Mol Sci. 2016. PMID: 26907255 Free PMC article. Review.
-
Generation of CD34+ cells from human embryonic stem cells using a clinically applicable methodology and engraftment in the fetal sheep model.Exp Hematol. 2013 Aug;41(8):749-758.e5. doi: 10.1016/j.exphem.2013.04.003. Epub 2013 Apr 20. Exp Hematol. 2013. PMID: 23612043 Free PMC article.
-
E, K, B5, B6, and B9 vitamins and their specific immunological effects evaluated by flow cytometry.Front Med (Lausanne). 2023 Jan 5;9:1089476. doi: 10.3389/fmed.2022.1089476. eCollection 2022. Front Med (Lausanne). 2023. PMID: 36687400 Free PMC article. Review.
-
The CD34 surface antigen is restricted to glucagon-expressing cells in the early developing bovine pancreas.Histochem Cell Biol. 2011 Jan;135(1):59-71. doi: 10.1007/s00418-010-0775-x. Epub 2011 Jan 4. Histochem Cell Biol. 2011. PMID: 21203769
-
In utero transplantation: Disparate ramifications.World J Stem Cells. 2013 Apr 26;5(2):43-52. doi: 10.4252/wjsc.v5.i2.43. World J Stem Cells. 2013. PMID: 23671718 Free PMC article.
References
-
- Hanafusa T, Miyazaki A, Miyagawa J, et al. Examination of islets in the pancreas biopsy specimens from newly diagnosed type 1 (insulin-dependent) diabetic patients. Diabetologia. 1990;33:105–111. - PubMed
-
- Butler AE, Janson J, Bonner-Weir S, Ritzel R, Rizza RA, Butler PC. {beta}-Cell Deficit and Increased {beta}-Cell Apoptosis in Humans With Type 2 Diabetes. Diabetes. 52:102–110. - PubMed
-
- Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science. 1998;282:1145–1147. 2003. - PubMed
-
- Kroon E, Martinson LA, Kadoya K, et al. Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat Biotechnol. 2008;26:443–452. - PubMed
-
- Brolen GK, Heins N, Edsbagge J, Semb H. Signals from the embryonic mouse pancreas induce differentiation of human embryonic stem cells into insulin-producing beta-cell-like cells. Diabetes. 2005;54:2867–2874. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
