

Mycobacterium tuberculosis MycP1 protease plays a dual role in regulation of ESX-1 secretion and virulence
- PMID: 20227664
- PMCID: PMC3121311
- DOI: 10.1016/j.chom.2010.02.006
Mycobacterium tuberculosis MycP1 protease plays a dual role in regulation of ESX-1 secretion and virulence
Abstract
Mycobacterium tuberculosis uses the ESX-1 secretion system to deliver virulence proteins during infection of host cells. Here we report a mechanism of posttranscriptional control of ESX-1 mediated by MycP1, a M. tuberculosis serine protease. We show that MycP1 is required for ESX-1 secretion but that, unexpectedly, genetic inactivation of MycP1 protease activity increases secretion of ESX-1 substrates. We demonstrate that EspB, an ESX-1 substrate required for secretion, is a target of MycP1 in vitro and in vivo. During macrophage infection, an inactive MycP1 protease mutant causes hyperactivation of ESX-1-stimulated innate signaling pathways. MycP1 is required for growth in mice during acute infection, while loss of its protease activity leads to attenuated virulence during chronic infection. As the key ESX-1 substrates ESAT-6 and CFP-10 are highly immunogenic, fine-tuning of their secretion by MycP1 may balance virulence and immune detection and be essential for successful maintenance of long-term M. tuberculosis infection.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
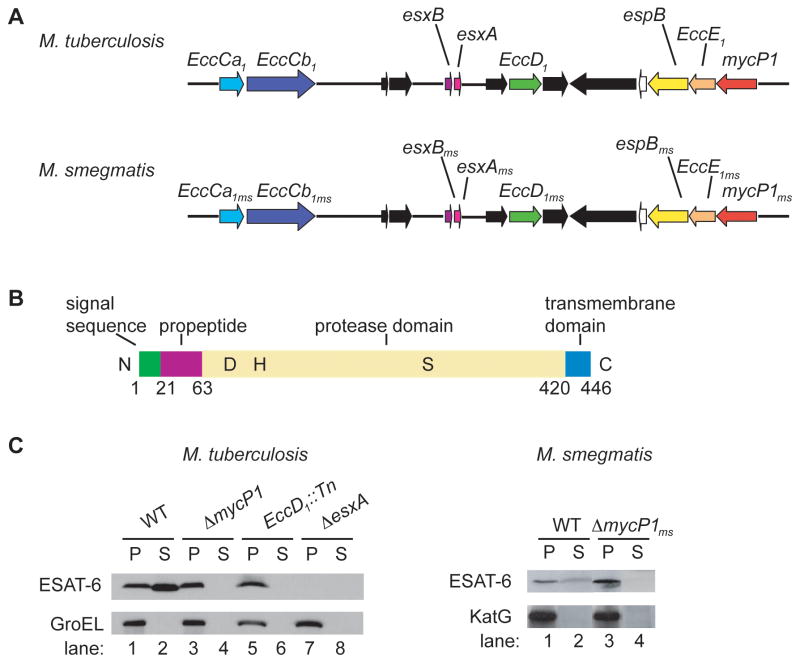
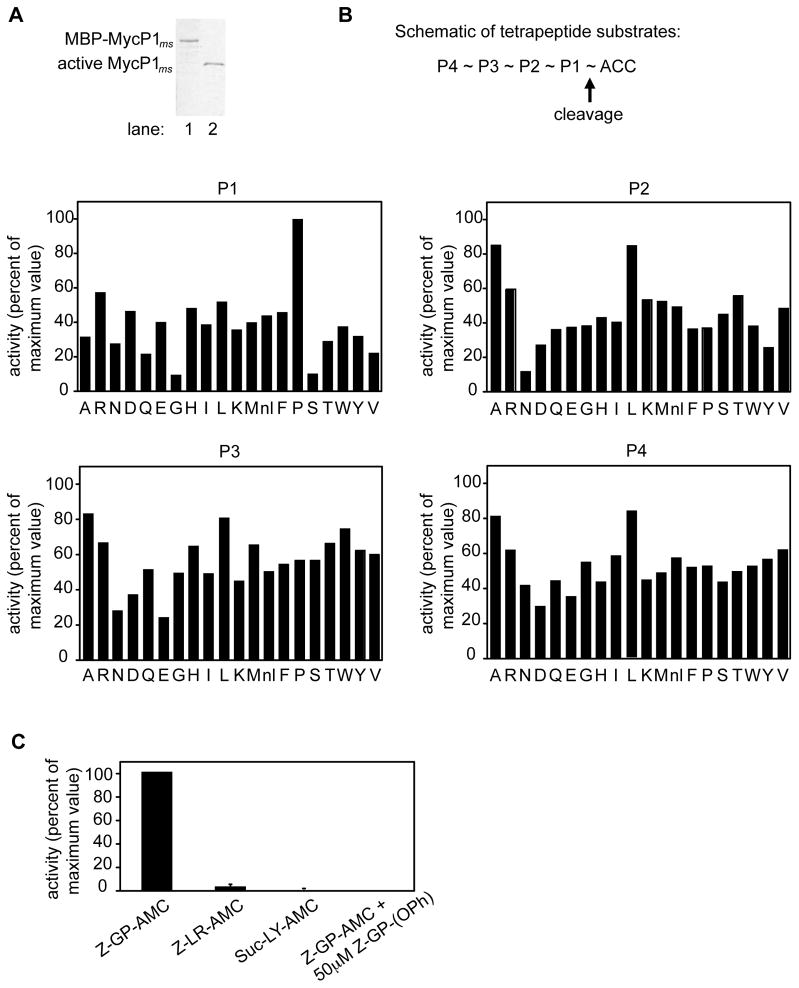
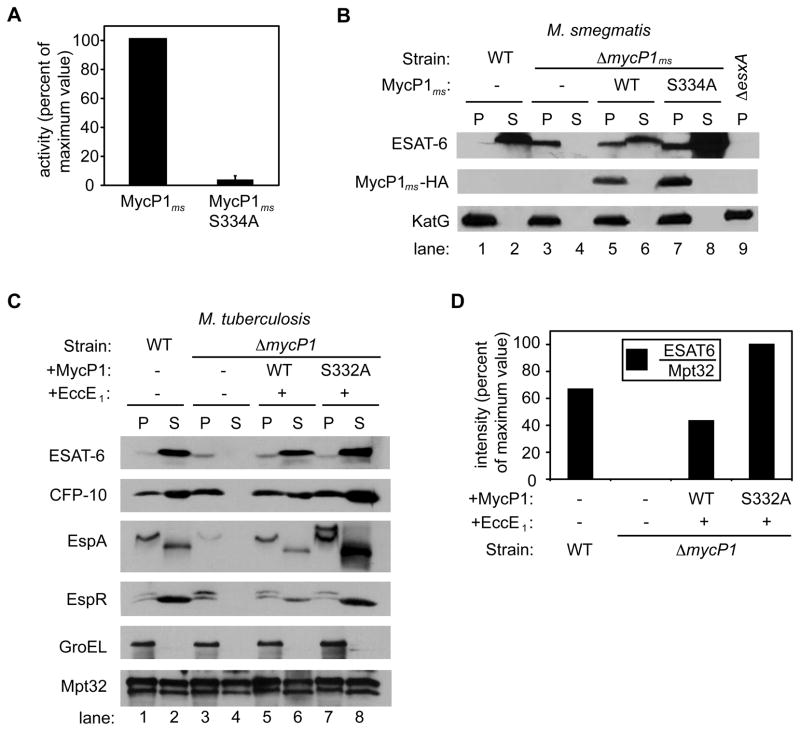
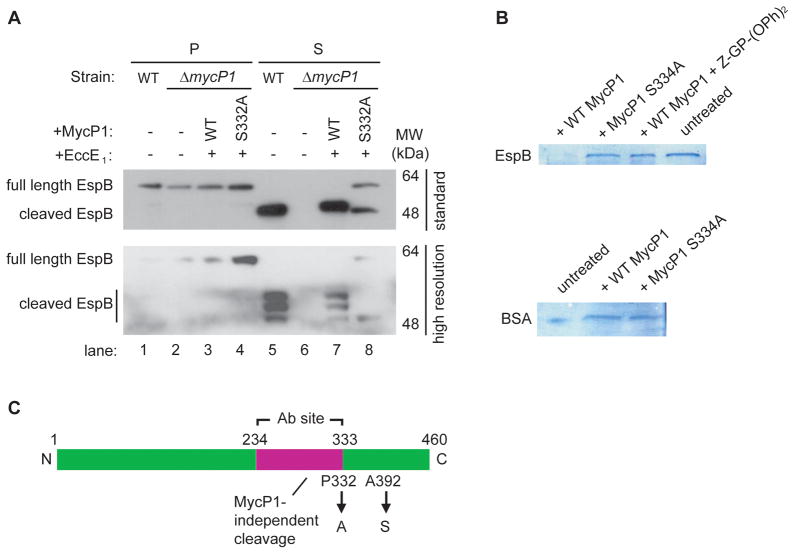
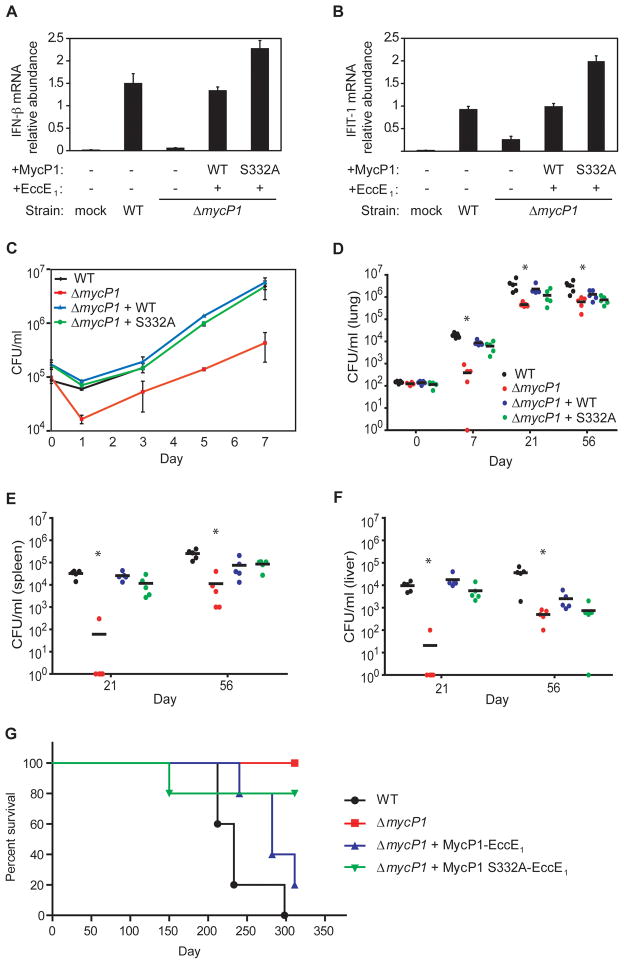
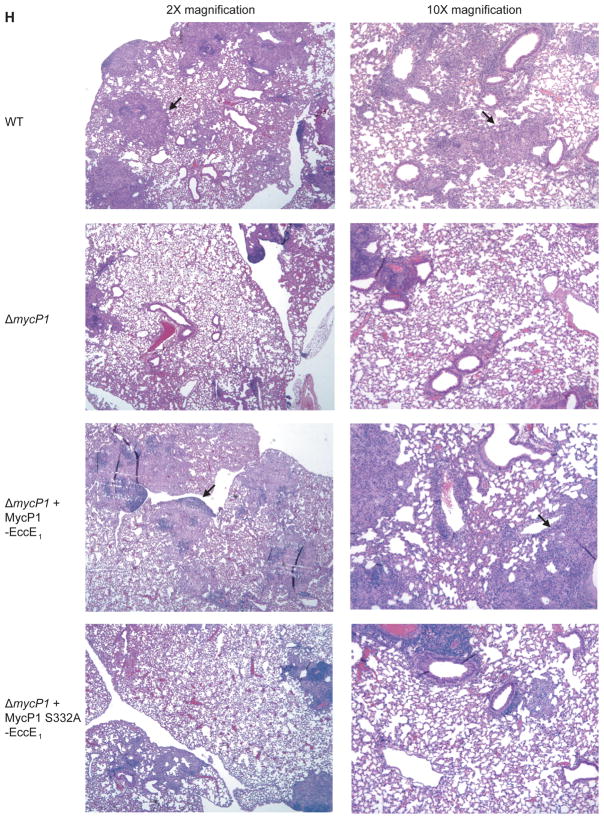
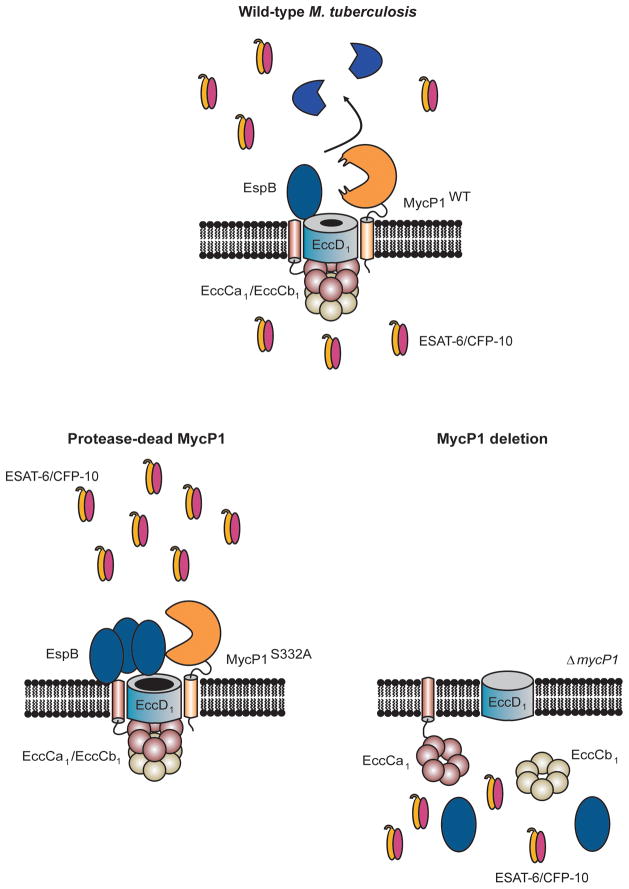
Comment in
-
The two faces of MycP1.Nat Rev Microbiol. 2010 May;8(5):314. doi: 10.1038/nrmicro2360. Nat Rev Microbiol. 2010. PMID: 21080601 No abstract available.
References
-
- Abdallah AM, Gey van Pittius NC, Champion PAD, Cox JS, Luirink J, Vandenbroucke-Grauls CMJE, Appelmelk BJ, Bitter W. Type VII secretion – mycobacteria show the way. Nature Reviews. 2007;5:883–891. - PubMed
-
- Bergeron F, Leduc R, Day R. Subtilase-like pro-protein convertases: from molecular specificity to therapeutic applications. J Mol Endocrinol. 2000;24:1–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
