

The Cellular lysine methyltransferase Set7/9-KMT7 binds HIV-1 TAR RNA, monomethylates the viral transactivator Tat, and enhances HIV transcription
- PMID: 20227666
- PMCID: PMC2844784
- DOI: 10.1016/j.chom.2010.02.005
The Cellular lysine methyltransferase Set7/9-KMT7 binds HIV-1 TAR RNA, monomethylates the viral transactivator Tat, and enhances HIV transcription
Abstract
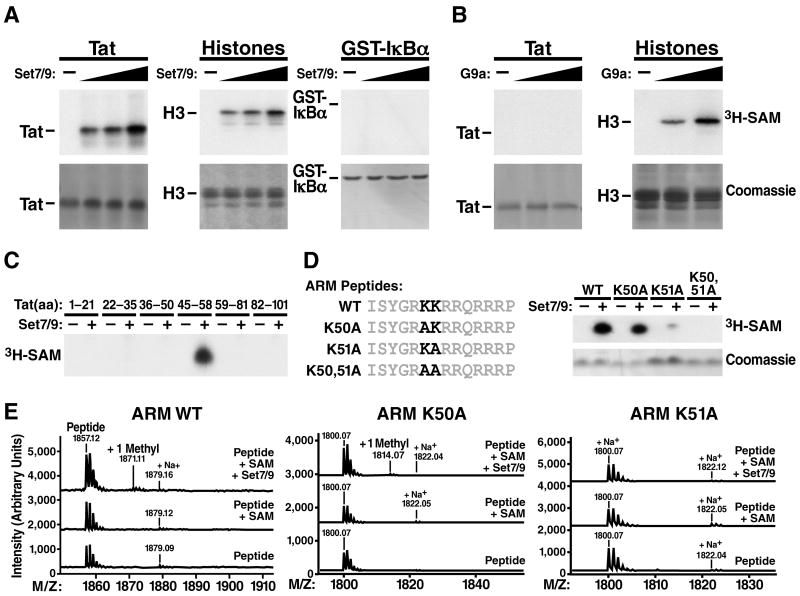
The Tat protein of HIV-1 plays an essential role in HIV gene expression by promoting efficient elongation of viral transcripts. Posttranslational modifications of Tat fine-tune interactions of Tat with cellular cofactors and TAR RNA, a stem-loop structure at the 5' ends of viral transcripts. Here, we identify the lysine methyltransferase Set7/9 (KMT7) as a coactivator of HIV transcription. Set7/9-KMT7 associates with the HIV promoter in vivo and monomethylates lysine 51, a highly conserved residue located in the RNA-binding domain of Tat. Knockdown of Set7/9-KMT7 suppresses Tat transactivation of the HIV promoter, but does not affect the transcriptional activity of methylation-deficient Tat (K51A). Set7/9-KMT7 binds TAR RNA by itself and in complex with Tat and the positive transcription elongation factor P-TEFb. Our findings uncover a positive role for Set7/9-KMT7 and Tat methylation during early steps of the Tat transactivation cycle.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures






Comment in
-
The HIV-1 Tat team gets bigger.Cell Host Microbe. 2010 Mar 18;7(3):179-81. doi: 10.1016/j.chom.2010.03.001. Cell Host Microbe. 2010. PMID: 20227660
References
-
- Anand K, Schulte A, Vogel-Bachmayr K, Scheffzek K, Geyer M. Structural insights into the cyclin T1-Tat-TAR RNA transcription activation complex from EIAV. Nat Struct Mol Biol. 2008;15:1287–1292. - PubMed
-
- Benkirane M, Chun RF, Xiao H, Ogryzko VV, Howard BH, Nakatani Y, Jeang KT. Activation of integrated provirus requires histone acetyltransferase. p300 and P/CAF are coactivators for HIV-1 Tat. J Biol Chem. 1998;273:24898–24905. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
