

Role of protein misfolding in DFNA9 hearing loss
- PMID: 20228067
- PMCID: PMC2865277
- DOI: 10.1074/jbc.M110.106724
Role of protein misfolding in DFNA9 hearing loss
Abstract
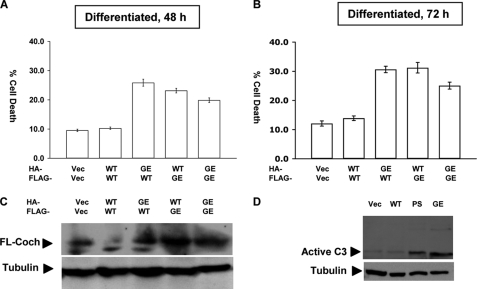
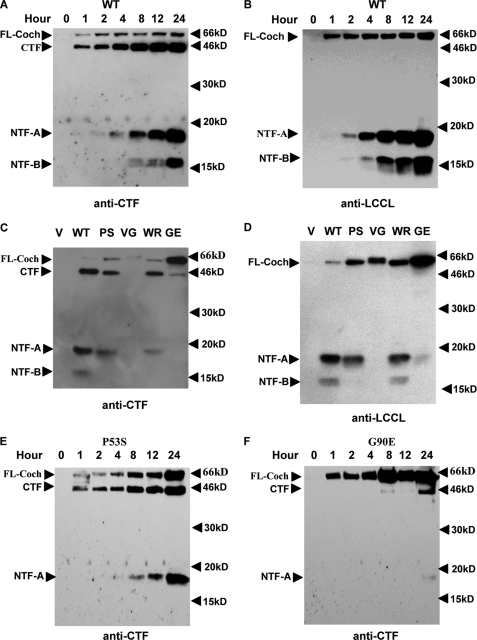
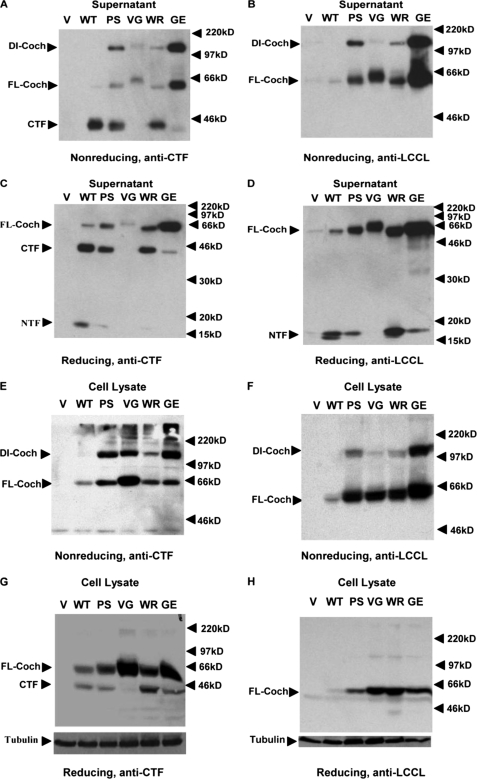
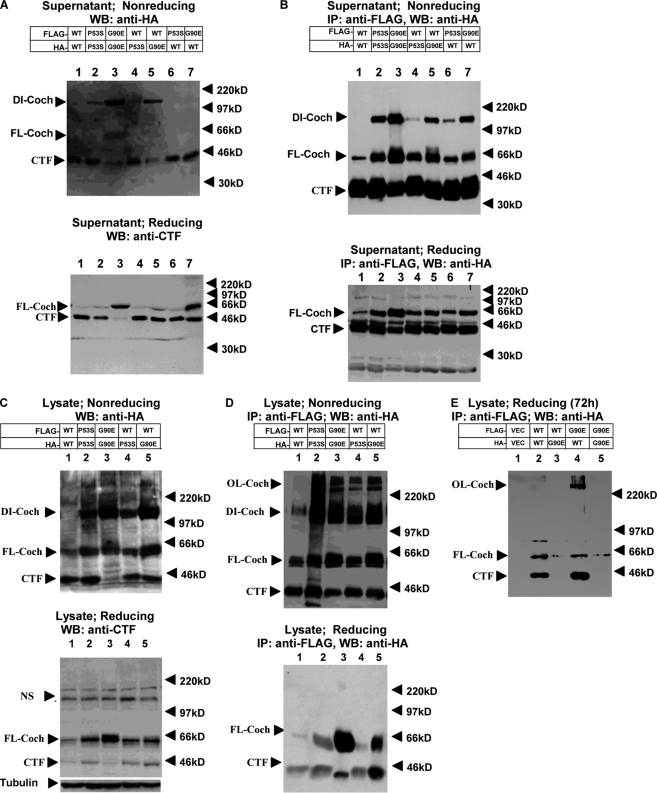
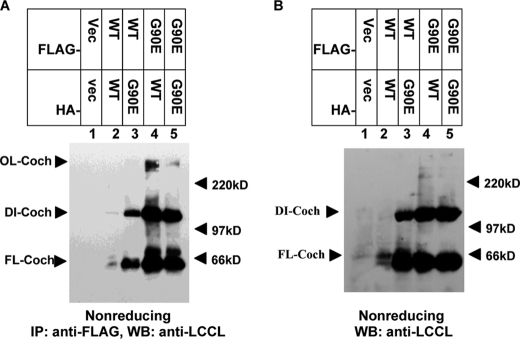
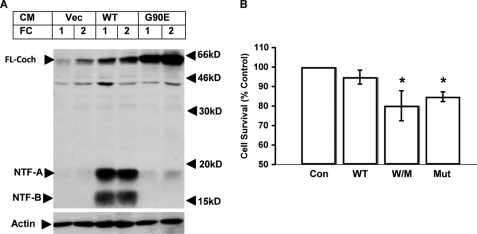
Mutations in the COCH (coagulation factor C homology) gene have been attributed to DFNA9 (deafness, autosomal-dominant 9), an autosomal-dominant non-syndromic hearing loss disorder. However, the mechanisms responsible for DFNA9 hearing loss remain unknown. Here, we demonstrate that mutant cochlin, the protein product of the COCH gene, forms a stable dimer that is sensitive to reducing agent. In contrast, wild-type (WT) cochlin may form only dimers transiently. Interestingly, the presence of mutant cochlin can stabilize WT cochlin in dimer conformation, providing a possible mechanism for the dominant nature of DFNA9 mutations. Furthermore, the expression of mutant cochlin eventually induces WT cochlin to form stable oligomers that are resistant to reducing agent. Finally, we show that mutant cochlin is cytotoxic in vitro and in vivo. Our study suggests a possible molecular mechanism underlying DFNA9 hearing loss and provides an in vitro model that may be used to explore protein-misfolding diseases in general.
Figures
References
-
- Bossy-Wetzel E., Schwarzenbacher R., Lipton S. A. (2004) Nat. Med. 10, (suppl.) S2–S9 - PubMed
-
- Robertson N. G., Lu L., Heller S., Merchant S. N., Eavey R. D., McKenna M., Nadol J. B., Jr., Miyamoto R. T., Linthicum F. H., Jr., Lubianca Neto J. F., Hudspeth A. J., Seidman C. E., Morton C. C., Seidman J. G. (1998) Nat. Genet. 20, 299–303 - PubMed
-
- Merchant S. N., Linthicum F. H., Nadol J. B., Jr. (2000) Adv. Otorhinolaryngol. 56, 212–217 - PubMed
-
- Khetarpal U., Schuknecht H. F., Gacek R. R., Holmes L. B. (1991) Arch. Otolaryngol. Head Neck Surg. 117, 1032–1042 - PubMed
-
- Robertson N. G., Skvorak A. B., Yin Y., Weremowicz S., Johnson K. R., Kovatch K. A., Battey J. F., Bieber F. R., Morton C. C. (1997) Genomics 46, 345–354 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
