

Boron transport in plants: co-ordinated regulation of transporters
- PMID: 20228086
- PMCID: PMC2887066
- DOI: 10.1093/aob/mcq044
Boron transport in plants: co-ordinated regulation of transporters
Abstract
Background: The essentiality of boron (B) for plant growth was established > 85 years ago. In the last decade, it has been revealed that one of the physiological roles of B is cross-linking the pectic polysaccharide rhamnogalacturonan II in primary cell walls. Borate cross-linking of pectic networks serves both for physical strength of cell walls and for cell adhesion. On the other hand, high concentrations of B are toxic to plant growth. To avoid deficiency and toxicity problems, it is important for plants to maintain their tissue B concentrations within an optimum range by regulating transport processes. Boron transport was long believed to be a passive, unregulated process, but the identification of B transporters has suggested that plants sense and respond to the B conditions and regulate transporters to maintain B homeostasis.
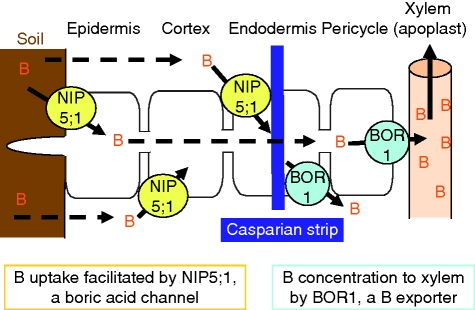
Scope: Transporters responsible for efficient B uptake by roots, xylem loading and B distribution among leaves have been described. These transporters are required under B limitation for efficient acquisition and utilization of B. Transporters important for tolerating high B levels in the environment have also been identified, and these transporters export B from roots back to the soil. Two types of transporters are involved in these processes: NIPs (nodulin-26-like intrinsic proteins), boric acid channels, and BORs, B exporters. It is demonstrated that the expression of genes encoding these transporters is finely regulated in response to B availability in the environment to ensure tissue B homeostasis. Furthermore, plants tolerant to stress produced by low B or high B in the environment can be generated through altered expression of these transporters.
Conclusions: The identification of the first B transporter led to the discovery that B transport was a process mediated not only by passive diffusion but also by transporters whose activity was regulated in response to B conditions. Now it is evident that plants sense internal and external B conditions and regulate B transport by modulating the expression and/or accumulation of these transporters. Results obtained in model plants are applicable to other plant species, and such knowledge may be useful in designing plants or crops tolerant to soils containing low or high B.
Figures
Similar articles
-
Regulation, Diversity and Evolution of Boron Transporters in Plants.Plant Cell Physiol. 2021 Sep 24;62(4):590-599. doi: 10.1093/pcp/pcab025. Plant Cell Physiol. 2021. PMID: 33570563 Review.
-
Boron transport mechanisms: collaboration of channels and transporters.Trends Plant Sci. 2008 Aug;13(8):451-7. doi: 10.1016/j.tplants.2008.05.007. Epub 2008 Jul 4. Trends Plant Sci. 2008. PMID: 18603465 Review.
-
Cell-type specificity of the expression of Os BOR1, a rice efflux boron transporter gene, is regulated in response to boron availability for efficient boron uptake and xylem loading.Plant Cell. 2007 Aug;19(8):2624-35. doi: 10.1105/tpc.106.049015. Epub 2007 Aug 3. Plant Cell. 2007. PMID: 17675406 Free PMC article.
-
Molecular mechanisms of boron transport in plants: involvement of Arabidopsis NIP5;1 and NIP6;1.Adv Exp Med Biol. 2010;679:83-96. doi: 10.1007/978-1-4419-6315-4_7. Adv Exp Med Biol. 2010. PMID: 20666226 Review.
-
Homeostasis of the structurally important micronutrients, B and Si.Curr Opin Plant Biol. 2009 Jun;12(3):307-11. doi: 10.1016/j.pbi.2009.04.007. Epub 2009 May 27. Curr Opin Plant Biol. 2009. PMID: 19481495 Review.
Cited by
-
Tetraploidy Enhances Boron-Excess Tolerance in Carrizo Citrange (Citrus sinensis L. Osb. × Poncirus trifoliata L. Raf.).Front Plant Sci. 2016 May 25;7:701. doi: 10.3389/fpls.2016.00701. eCollection 2016. Front Plant Sci. 2016. PMID: 27252717 Free PMC article.
-
Roles of BOR2, a boron exporter, in cross linking of rhamnogalacturonan II and root elongation under boron limitation in Arabidopsis.Plant Physiol. 2013 Dec;163(4):1699-709. doi: 10.1104/pp.113.225995. Epub 2013 Oct 10. Plant Physiol. 2013. PMID: 24114060 Free PMC article.
-
Long-term changes in soil biological activity and other properties of raised beds in Longan orchards.PeerJ. 2024 Nov 6;12:e18396. doi: 10.7717/peerj.18396. eCollection 2024. PeerJ. 2024. PMID: 39677944 Free PMC article.
-
Mathematical modeling and experimental validation of the spatial distribution of boron in the root of Arabidopsis thaliana identify high boron accumulation in the tip and predict a distinct root tip uptake function.Plant Cell Physiol. 2015 Apr;56(4):620-30. doi: 10.1093/pcp/pcv016. Epub 2015 Feb 9. Plant Cell Physiol. 2015. PMID: 25670713 Free PMC article.
-
Proteomic and Genomic Studies of Micronutrient Deficiency and Toxicity in Plants.Plants (Basel). 2022 Sep 16;11(18):2424. doi: 10.3390/plants11182424. Plants (Basel). 2022. PMID: 36145825 Free PMC article. Review.
References
-
- Ahn JW, Verma R, Kim M, et al. Depletion of UDP-d-apiose/UDP-d-xylose synthases results in rhamnogalacturonan-II deficiency, cell wall thickening, and cell death in higher plants. Journal of Biological Chemistry. 2006;281:13708–13716. - PubMed
-
- Amin SA, Kupper FC, Green DH, Harris WR, Carrano CJ. Boron binding by a siderophore isolated from marine bacteria associated with the toxic dinoflagellate Gymnodinium catenatum. Journal of the American Chemical Society. 2007;129:478–479. - PubMed
-
- Bolanos L, Lukaszewski K, Bonilla I, Blevins D. Why boron? Plant Physiology and Biochemistry. 2004;42:907–912. - PubMed
