

Rapid degeneration and regeneration of the zebrafish olfactory epithelium after triton X-100 application
- PMID: 20228140
- PMCID: PMC2871777
- DOI: 10.1093/chemse/bjq019
Rapid degeneration and regeneration of the zebrafish olfactory epithelium after triton X-100 application
Abstract
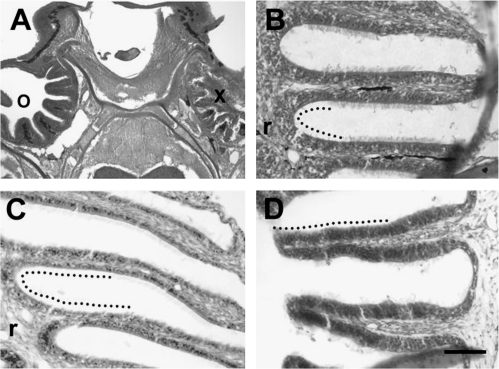
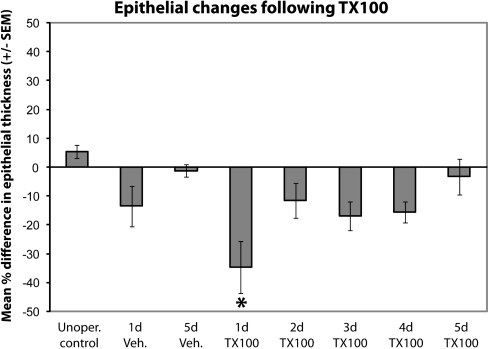
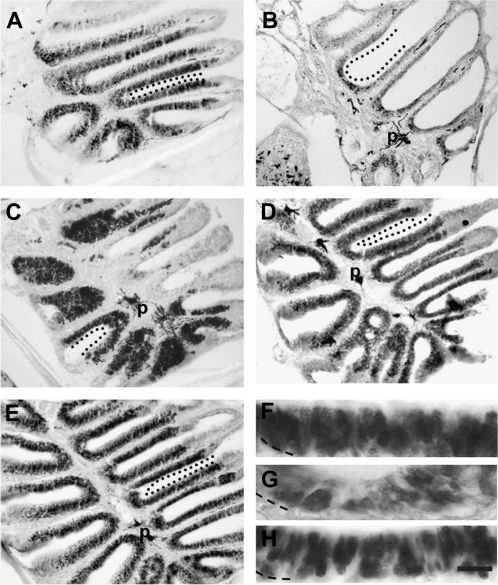
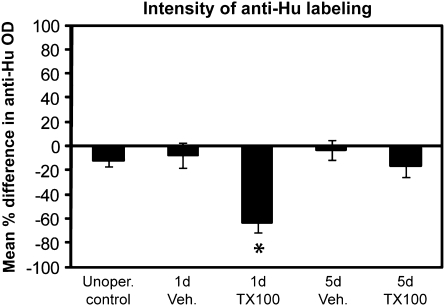
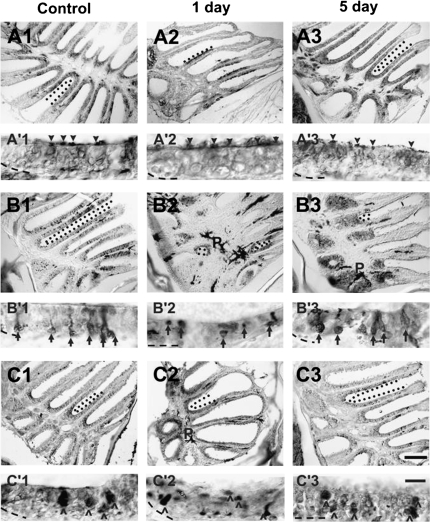
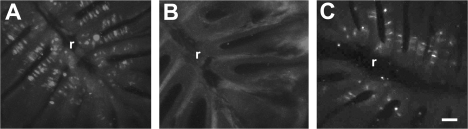
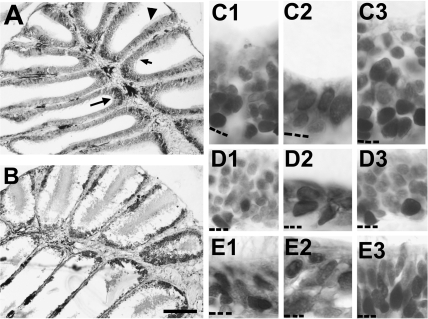
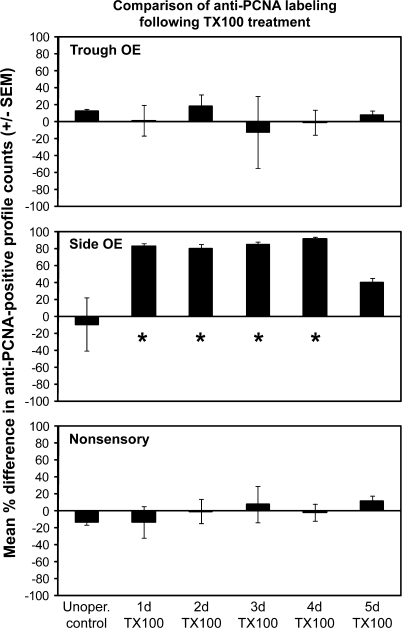
The effects of Triton X-100 on the olfactory epithelium (OE) of adult zebrafish were examined to study neuronal turnover in this model system. Fish were killed at various time points after detergent application and stained with hematoxylin and eosin to examine olfactory structures, immunocytochemistry to examine cell types, or DiI to examine connections to the olfactory bulb. A significant decrease in epithelial thickness of treated sides was observed 1-day posttreatment. Epithelium thickness recovered by 5 days. The most significant reduction in the OE following Triton X-100 treatment corresponded to the region of supporting cells and mature olfactory sensory neurons. Labeling for all neurons with anti-Hu and for the 3 sensory neuron subtypes of the zebrafish OE (ciliated, microvillous, and crypt neurons) diminished 1 day after lesion and returned by 5 days posttreatment. Retrograde labeling from the olfactory bulb showed that the majority of mature olfactory sensory neurons disappeared in 1 day and reappeared by 5 days after treatment. Anti-proliferating cell nuclear antigen was used to show mitotic activity, and after chemical lesion, there was an increase in proliferation in specific regions of the OE. Thus, chemical ablation causes temporary reduction with swift regeneration of the OE occurring within a week.
Figures
Similar articles
-
Olfactory bulb recovery following reversible deafferentation with repeated detergent application in the adult zebrafish.Neuroscience. 2011 Nov 24;196:276-84. doi: 10.1016/j.neuroscience.2011.09.005. Epub 2011 Sep 10. Neuroscience. 2011. PMID: 21933699
-
Plasticity of glomeruli and olfactory-mediated behavior in zebrafish following detergent lesioning of the olfactory epithelium.Neuroscience. 2015 Jan 22;284:622-631. doi: 10.1016/j.neuroscience.2014.10.036. Epub 2014 Oct 30. Neuroscience. 2015. PMID: 25450960 Free PMC article.
-
Differential copper-induced death and regeneration of olfactory sensory neuron populations and neurobehavioral function in larval zebrafish.Neurotoxicology. 2018 Dec;69:141-151. doi: 10.1016/j.neuro.2018.10.002. Epub 2018 Oct 4. Neurotoxicology. 2018. PMID: 30292653 Free PMC article.
-
Neurogenesis and cell death in olfactory epithelium.J Neurobiol. 1996 May;30(1):67-81. doi: 10.1002/(SICI)1097-4695(199605)30:1<67::AID-NEU7>3.0.CO;2-E. J Neurobiol. 1996. PMID: 8727984 Review.
-
Apoptosis in the mature and developing olfactory neuroepithelium.Microsc Res Tech. 2002 Aug 1;58(3):204-15. doi: 10.1002/jemt.10150. Microsc Res Tech. 2002. PMID: 12203699 Review.
Cited by
-
The nitroreductase system of inducible targeted ablation facilitates cell-specific regenerative studies in zebrafish.Methods. 2013 Aug 15;62(3):232-40. doi: 10.1016/j.ymeth.2013.03.017. Epub 2013 Mar 27. Methods. 2013. PMID: 23542552 Free PMC article.
-
A lifetime of neurogenesis in the olfactory system.Front Neurosci. 2014 Jun 26;8:182. doi: 10.3389/fnins.2014.00182. eCollection 2014. Front Neurosci. 2014. PMID: 25018692 Free PMC article. Review.
-
Activity-dependent formation of the topographic map and the critical period in the development of mammalian olfactory system.Genesis. 2024 Feb;62(1):e23586. doi: 10.1002/dvg.23586. Genesis. 2024. PMID: 38593162 Free PMC article. Review.
-
The Olfactory System of Zebrafish as a Model for the Study of Neurotoxicity and Injury: Implications for Neuroplasticity and Disease.Int J Mol Sci. 2019 Apr 2;20(7):1639. doi: 10.3390/ijms20071639. Int J Mol Sci. 2019. PMID: 30986990 Free PMC article. Review.
-
The Olfactory Organ Is a Unique Site for Neutrophils in the Brain.Front Immunol. 2022 May 27;13:881702. doi: 10.3389/fimmu.2022.881702. eCollection 2022. Front Immunol. 2022. PMID: 35693773 Free PMC article.
References
-
- Adamek GD, Gesteland RC, Mair RG, Oakley B. Transduction physiology of olfactory receptor cilia. Brain Res. 1984;310:87–97. - PubMed
-
- Adolf B, Chapouton P, Lam CS, Topp S. Conserved and acquired features of adult neurogenesis in the zebrafish telencephalon. Dev Biol. 2006;295:278–293. - PubMed
-
- Barth AL, Justice NJ, Ngai J. Asynchronous onset of odorant receptor expression in the developing zebrafish olfactory system. Neuron. 1996;16:23–34. - PubMed
-
- Becker CG, Becker T. Adult zebrafish as a model for successful central nervous system regeneration. Restor Neurol Neurosci. 2008;26:71–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
