

Neuromechanical simulation of the locust jump
- PMID: 20228342
- PMCID: PMC2837733
- DOI: 10.1242/jeb.034678
Neuromechanical simulation of the locust jump
Abstract
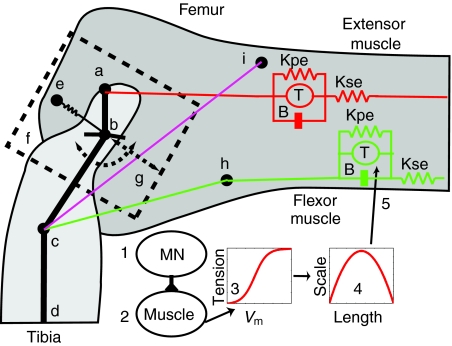
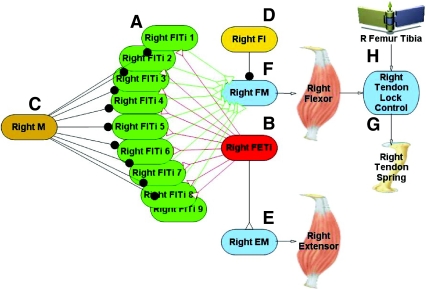
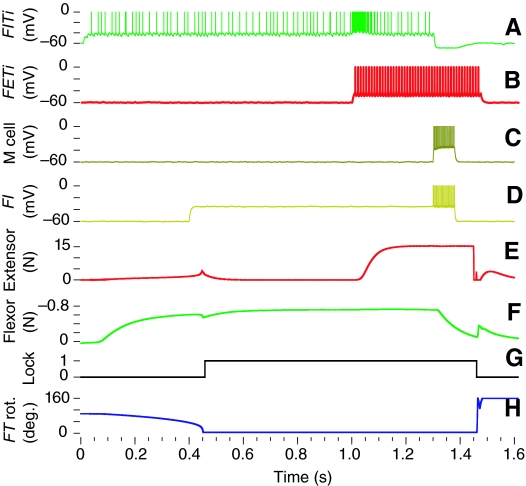
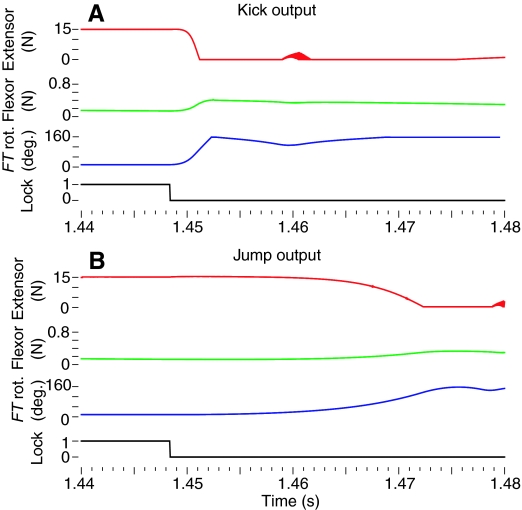
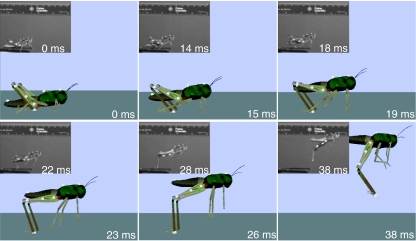
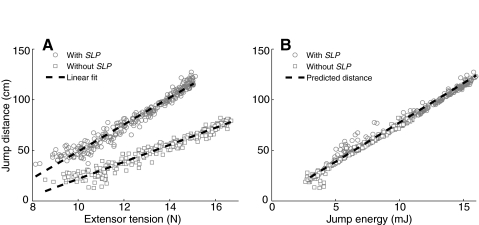
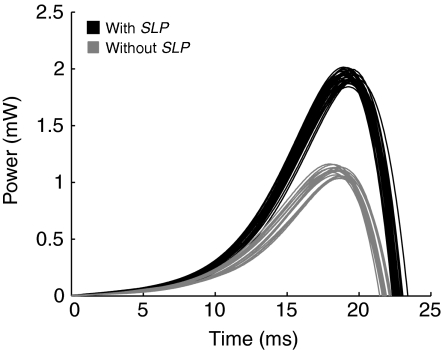
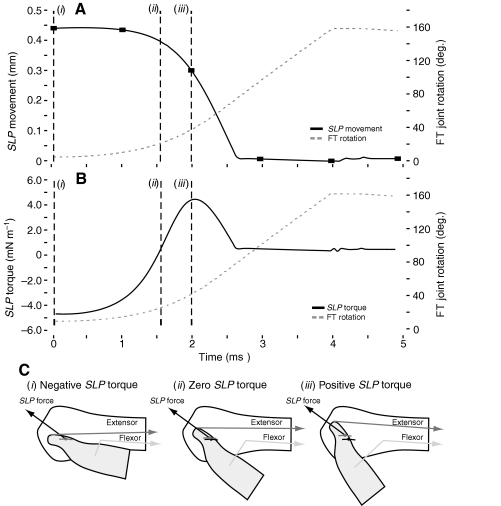
The neural circuitry and biomechanics of kicking in locusts have been studied to understand their roles in the control of both kicking and jumping. It has been hypothesized that the same neural circuit and biomechanics governed both behaviors but this hypothesis was not testable with current technology. We built a neuromechanical model to test this and to gain a better understanding of the role of the semi-lunar process (SLP) in jump dynamics. The jumping and kicking behaviors of the model were tested by comparing them with a variety of published data, and were found to reproduce the results from live animals. This confirmed that the kick neural circuitry can produce the jump behavior. The SLP is a set of highly sclerotized bands of cuticle that can be bent to store energy for use during kicking and jumping. It has not been possible to directly test the effects of the SLP on jump performance because it is an integral part of the joint, and attempts to remove its influence prevent the locust from being able to jump. Simulations demonstrated that the SLP can significantly increase jump distance, power, total energy and duration of the jump impulse. In addition, the geometry of the joint enables the SLP force to assist leg flexion when the leg is flexed, and to assist extension once the leg has begun to extend.
Figures
Similar articles
-
Locusts use a composite of resilin and hard cuticle as an energy store for jumping and kicking.J Exp Biol. 2012 Oct 1;215(Pt 19):3501-12. doi: 10.1242/jeb.071993. Epub 2012 Jun 12. J Exp Biol. 2012. PMID: 22693029
-
Jumping and kicking in the false stick insect Prosarthria teretrirostris: kinematics and motor control.J Exp Biol. 2002 Jun;205(Pt 11):1519-30. doi: 10.1242/jeb.205.11.1519. J Exp Biol. 2002. PMID: 12000798
-
The locust jump. II. Neural circuits of the motor programme.J Exp Biol. 1977 Feb;66(1):221-41. doi: 10.1242/jeb.66.1.221. J Exp Biol. 1977. PMID: 870600
-
When skeletons are geared for speed: the morphology, biomechanics, and energetics of rapid animal motion.Integr Comp Biol. 2012 Nov;52(5):588-96. doi: 10.1093/icb/ics111. Epub 2012 Sep 2. Integr Comp Biol. 2012. PMID: 22945756 Review.
-
Muscle coordination of movement: a perspective.J Biomech. 1993;26 Suppl 1:109-24. doi: 10.1016/0021-9290(93)90083-q. J Biomech. 1993. PMID: 8505346 Review.
Cited by
-
Passive joint forces are tuned to limb use in insects and drive movements without motor activity.Curr Biol. 2013 Aug 5;23(15):1418-26. doi: 10.1016/j.cub.2013.06.024. Epub 2013 Jul 18. Curr Biol. 2013. PMID: 23871240 Free PMC article.
-
A Survey of Bioinspired Jumping Robot: Takeoff, Air Posture Adjustment, and Landing Buffer.Appl Bionics Biomech. 2017;2017:4780160. doi: 10.1155/2017/4780160. Epub 2017 Sep 14. Appl Bionics Biomech. 2017. PMID: 29311756 Free PMC article. Review.
-
A physical model of mantis shrimp for exploring the dynamics of ultrafast systems.Proc Natl Acad Sci U S A. 2021 Aug 17;118(33):e2026833118. doi: 10.1073/pnas.2026833118. Proc Natl Acad Sci U S A. 2021. PMID: 34389671 Free PMC article.
-
NeuroMechFly, a neuromechanical model of adult Drosophila melanogaster.Nat Methods. 2022 May;19(5):620-627. doi: 10.1038/s41592-022-01466-7. Epub 2022 May 11. Nat Methods. 2022. PMID: 35545713
-
Latch-based control of energy output in spring actuated systems.J R Soc Interface. 2020 Jul;17(168):20200070. doi: 10.1098/rsif.2020.0070. Epub 2020 Jul 22. J R Soc Interface. 2020. PMID: 32693743 Free PMC article.
References
-
- Bennet-Clark H. C. (1975). The energetics of the jump of the locust Schistocerca gregaria. J. Exp. Biol. 63, 53-83 - PubMed
-
- Bennet-Clark H. C., Lucey E. C. (1967). The jump of the flea: a study of the energetics and a model of the mechanism. J. Exp. Biol. 47, 59-67 - PubMed
-
- Burrows M. (1995). Motor patterns during kicking movements in the locust. J. Comp. Physiol. A 176, 289-305 - PubMed
-
- Burrows M. (1996). The Neurobiology of an Insect Brain Oxford: Oxford University Press;
-
- Burrows M. (2003). Biomechanics: froghopper insects leap to new heights. Nature 424, 509 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
