

Hsk1- and SCF(Pof3)-dependent proteolysis of S. pombe Ams2 ensures histone homeostasis and centromere function
- PMID: 20230746
- PMCID: PMC2880248
- DOI: 10.1016/j.devcel.2009.12.024
Hsk1- and SCF(Pof3)-dependent proteolysis of S. pombe Ams2 ensures histone homeostasis and centromere function
Abstract
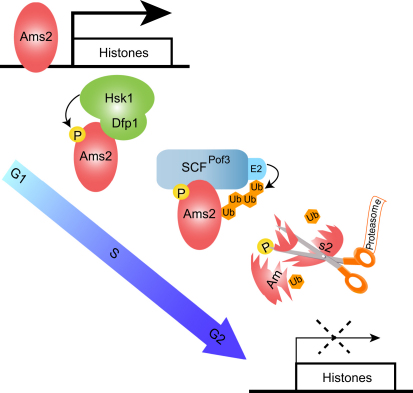
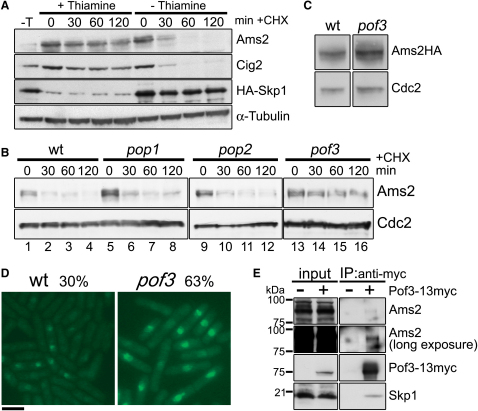
Schizosaccharomyces pombe GATA factor Ams2 is responsible for cell cycle-dependent transcriptional activation of all the core histone genes peaking at G1/S phase. Intriguingly, its own protein level also fluctuates concurrently. Here, we show that Ams2 is ubiquitylated and degraded through the SCF (Skp1-Cdc53/Cullin-1-F-box) ubiquitin ligase, in which F box protein Pof3 binds this protein. Ams2 is phosphorylated at multiple sites, which is required for SCF(Pof3)-dependent proteolysis. Hsk1/Cdc7 kinase physically associates with and phosphorylates Ams2. Even mild overexpression of Ams2 induces constitutive histone expression and chromosome instability, and its toxicity is exaggerated when Hsk1 function is compromised. This is partly attributable to abnormal incorporation of canonical H3 into the central CENP-A/Cnp1-rich centromere, thereby reversing specific chromatin structures to apparently normal nucleosomes. We propose that Hsk1 plays a vital role during post S phase in genome stability via SCF(Pof3)-mediated degradation of Ams2, thereby maintaining centromere integrity.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Terminating histone synthesis to preserve centromere integrity.Dev Cell. 2010 Mar 16;18(3):335-6. doi: 10.1016/j.devcel.2010.03.004. Dev Cell. 2010. PMID: 20230740
Similar articles
-
Anaphase-promoting complex/cyclosome-mediated proteolysis of Ams2 in the G1 phase ensures the coupling of histone gene expression to DNA replication in fission yeast.J Biol Chem. 2013 Jan 11;288(2):928-37. doi: 10.1074/jbc.M112.410241. Epub 2012 Nov 29. J Biol Chem. 2013. PMID: 23195958 Free PMC article.
-
A cell cycle-regulated GATA factor promotes centromeric localization of CENP-A in fission yeast.Mol Cell. 2003 Jan;11(1):175-87. doi: 10.1016/s1097-2765(03)00011-x. Mol Cell. 2003. PMID: 12535531
-
Fission yeast F-box protein Pof3 is required for genome integrity and telomere function.Mol Biol Cell. 2002 Jan;13(1):211-24. doi: 10.1091/mbc.01-07-0333. Mol Biol Cell. 2002. PMID: 11809834 Free PMC article.
-
Two distinct pathways responsible for the loading of CENP-A to centromeres in the fission yeast cell cycle.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):595-606; discussion 606-7. doi: 10.1098/rstb.2004.1614. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897182 Free PMC article. Review.
-
Does a GATA factor make the bed for centromeric nucleosomes?Cell Cycle. 2003 Jul-Aug;2(4):277-8. Cell Cycle. 2003. PMID: 12851470 Review.
Cited by
-
Shaping the landscape: mechanistic consequences of ubiquitin modification of chromatin.EMBO Rep. 2012 Jun 29;13(7):619-30. doi: 10.1038/embor.2012.78. EMBO Rep. 2012. PMID: 22688965 Free PMC article. Review.
-
New insights into donor directionality of mating-type switching in Schizosaccharomyces pombe.PLoS Genet. 2018 May 31;14(5):e1007424. doi: 10.1371/journal.pgen.1007424. eCollection 2018 May. PLoS Genet. 2018. PMID: 29852001 Free PMC article.
-
Identification of Genes Encoding CENP-A and Heterochromatin Protein 1 of Lipomyces starkeyi and Functional Analysis Using Schizosaccharomyces pombe.Genes (Basel). 2020 Jul 8;11(7):769. doi: 10.3390/genes11070769. Genes (Basel). 2020. PMID: 32650514 Free PMC article.
-
Histone H2B Ubiquitylation Regulates Histone Gene Expression by Suppressing Antisense Transcription in Fission Yeast.Genetics. 2019 Sep;213(1):161-172. doi: 10.1534/genetics.119.302499. Epub 2019 Jul 25. Genetics. 2019. PMID: 31345994 Free PMC article.
-
Rad51-Rad52 mediated maintenance of centromeric chromatin in Candida albicans.PLoS Genet. 2014 Apr 24;10(4):e1004344. doi: 10.1371/journal.pgen.1004344. eCollection 2014 Apr. PLoS Genet. 2014. PMID: 24762765 Free PMC article.
References
-
- Bailis J.M., Bernard P., Antonelli R., Allshire R.C., Forsburg S.L. Hsk1-Dfp1 is required for heterochromatin-mediated cohesion at centromeres. Nat. Cell Biol. 2003;5:1111–1116. - PubMed
-
- Brown G.W., Kelly T.J. Purification of Hsk1, a minichromosome maintenance protein kinase from fission yeast. J. Biol. Chem. 1998;273:22083–22090. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
