

Sorting of the Alzheimer's disease amyloid precursor protein mediated by the AP-4 complex
- PMID: 20230749
- PMCID: PMC2841041
- DOI: 10.1016/j.devcel.2010.01.015
Sorting of the Alzheimer's disease amyloid precursor protein mediated by the AP-4 complex
Abstract
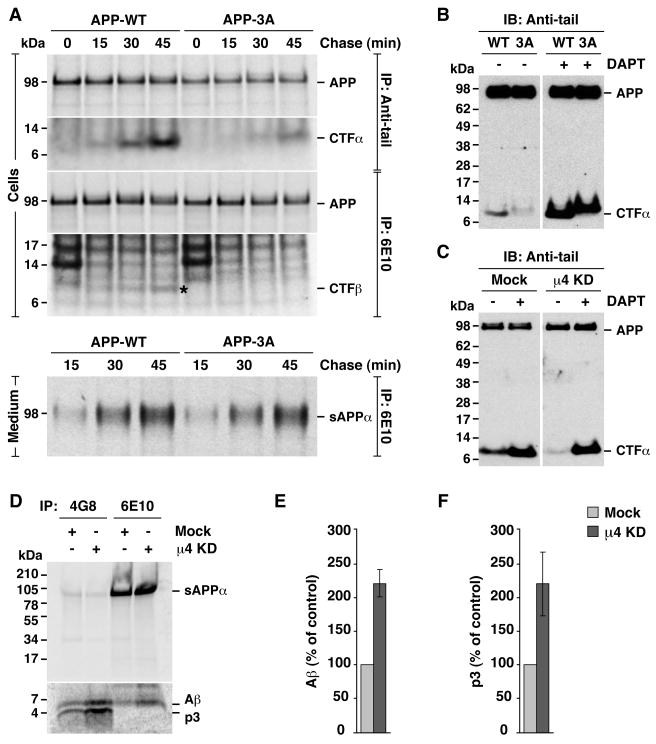
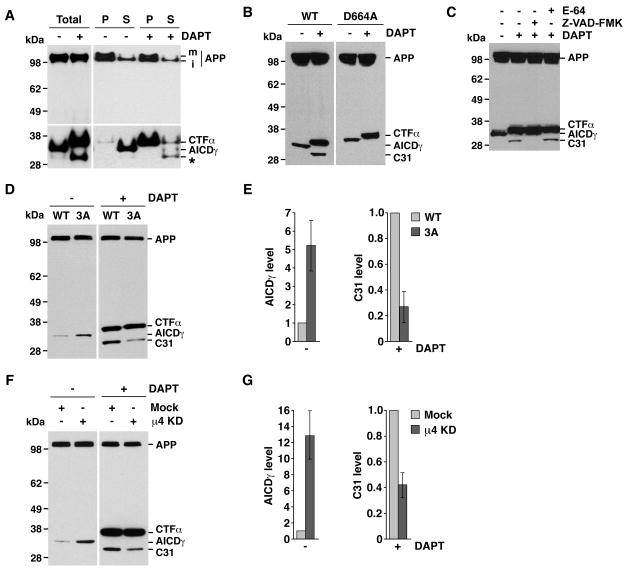
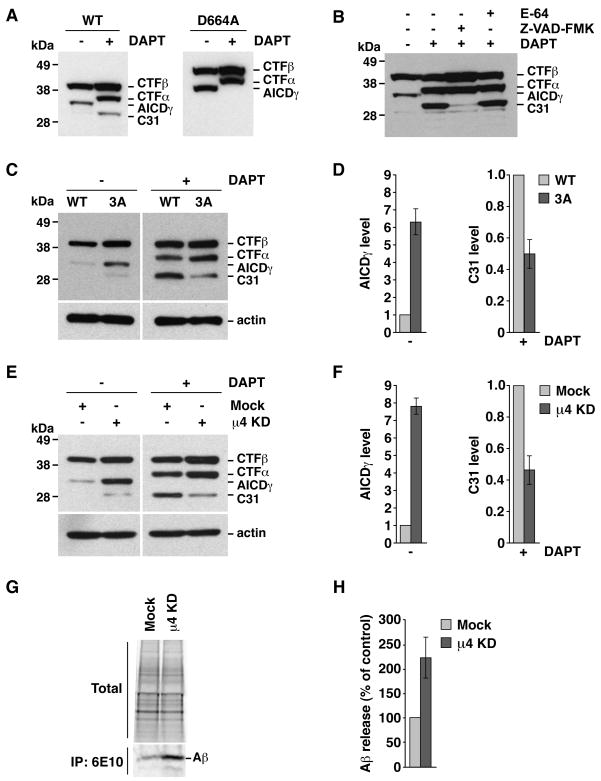
Adaptor protein 4 (AP-4) is the most recently discovered and least well-characterized member of the family of heterotetrameric adaptor protein (AP) complexes that mediate sorting of transmembrane cargo in post-Golgi compartments. Herein, we report the interaction of an YKFFE sequence from the cytosolic tail of the Alzheimer's disease amyloid precursor protein (APP) with the mu4 subunit of AP-4. Biochemical and X-ray crystallographic analyses reveal that the properties of the APP sequence and the location of the binding site on mu4 are distinct from those of other signal-adaptor interactions. Disruption of the APP-AP-4 interaction decreases localization of APP to endosomes and enhances gamma-secretase-catalyzed cleavage of APP to the pathogenic amyloid-beta peptide. These findings demonstrate that APP and AP-4 engage in a distinct type of signal-adaptor interaction that mediates transport of APP from the trans-Golgi network (TGN) to endosomes, thereby reducing amyloidogenic processing of the protein.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Structural and functional characterization of cargo-binding sites on the μ4-subunit of adaptor protein complex 4.PLoS One. 2014 Feb 3;9(2):e88147. doi: 10.1371/journal.pone.0088147. eCollection 2014. PLoS One. 2014. PMID: 24498434 Free PMC article.
-
Adaptor protein 2-mediated endocytosis of the β-secretase BACE1 is dispensable for amyloid precursor protein processing.Mol Biol Cell. 2012 Jun;23(12):2339-51. doi: 10.1091/mbc.E11-11-0944. Epub 2012 May 2. Mol Biol Cell. 2012. PMID: 22553349 Free PMC article.
-
LDLR-related protein 10 (LRP10) regulates amyloid precursor protein (APP) trafficking and processing: evidence for a role in Alzheimer's disease.Mol Neurodegener. 2012 Jun 26;7:31. doi: 10.1186/1750-1326-7-31. Mol Neurodegener. 2012. PMID: 22734645 Free PMC article.
-
Retromers in Alzheimer's disease.Neurodegener Dis. 2012;10(1-4):116-21. doi: 10.1159/000335910. Epub 2012 Mar 3. Neurodegener Dis. 2012. PMID: 22398391 Review.
-
Sorting receptor SORLA--a trafficking path to avoid Alzheimer disease.J Cell Sci. 2013 Jul 1;126(Pt 13):2751-60. doi: 10.1242/jcs.125393. Epub 2013 Jun 26. J Cell Sci. 2013. PMID: 23813966 Review.
Cited by
-
Role of the epithelial cell-specific clathrin adaptor complex AP-1B in cell polarity.Cell Logist. 2015 Jul 30;5(2):e1074331. doi: 10.1080/21592799.2015.1074331. eCollection 2015 Apr-Jun. Cell Logist. 2015. PMID: 27057418 Free PMC article. Review.
-
PAT1 inversely regulates the surface Amyloid Precursor Protein level in mouse primary neurons.BMC Neurosci. 2015 Mar 7;16:10. doi: 10.1186/s12868-015-0152-8. BMC Neurosci. 2015. PMID: 25880931 Free PMC article.
-
Calsyntenin-1 mediates axonal transport of the amyloid precursor protein and regulates Aβ production.Hum Mol Genet. 2012 Jul 1;21(13):2845-54. doi: 10.1093/hmg/dds109. Epub 2012 Mar 20. Hum Mol Genet. 2012. PMID: 22434822 Free PMC article.
-
Integrating structural and evolutionary data to interpret variation and pathogenicity in adapter protein complex 4.Protein Sci. 2020 Jun;29(6):1535-1549. doi: 10.1002/pro.3870. Epub 2020 Apr 25. Protein Sci. 2020. PMID: 32285480 Free PMC article.
-
GGA1 regulates signal-dependent sorting of BACE1 to recycling endosomes, which moderates Aβ production.Mol Biol Cell. 2018 Jan 15;29(2):191-208. doi: 10.1091/mbc.E17-05-0270. Epub 2017 Nov 15. Mol Biol Cell. 2018. PMID: 29142073 Free PMC article.
References
-
- Aguilar RC, Boehm M, Gorshkova I, Crouch RJ, Tomita K, Saito T, Ohno H, Bonifacino JS. Signal-binding specificity of the mu4 subunit of the adaptor protein complex AP-4. J Biol Chem. 2001;276:13145–13152. - PubMed
-
- Anliker B, Muller U. The functions of mammalian amyloid precursor protein and related amyloid precursor-like proteins. Neurodegener Dis. 2006;3:239–246. - PubMed
-
- Baulac S, LaVoie MJ, Kimberly WT, Strahle J, Wolfe MS, Selkoe DJ, Xia W. Functional gamma-secretase complex assembly in Golgi/trans-Golgi network: interactions among presenilin, nicastrin, Aph1, Pen-2, and gamma-secretase substrates. Neurobiol Dis. 2003;14:194–204. - PubMed
-
- Bonifacino JS, Traub LM. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu Rev Biochem. 2003;72:395–447. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
