

Separable functions of the fission yeast Spt5 carboxyl-terminal domain (CTD) in capping enzyme binding and transcription elongation overlap with those of the RNA polymerase II CTD
- PMID: 20231361
- PMCID: PMC2863715
- DOI: 10.1128/MCB.00116-10
Separable functions of the fission yeast Spt5 carboxyl-terminal domain (CTD) in capping enzyme binding and transcription elongation overlap with those of the RNA polymerase II CTD
Abstract
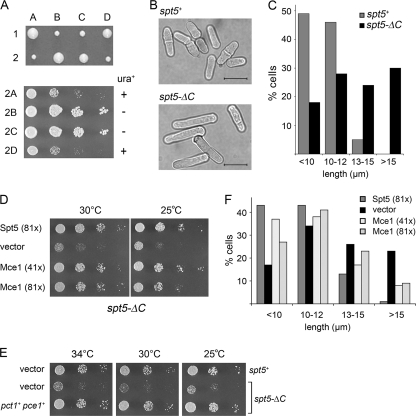
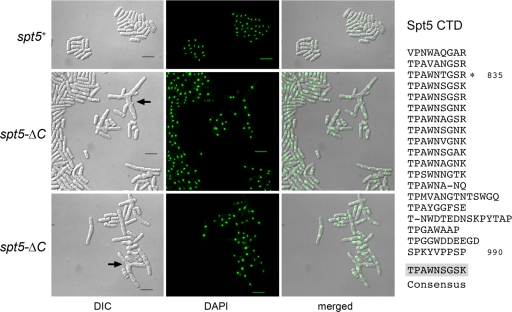
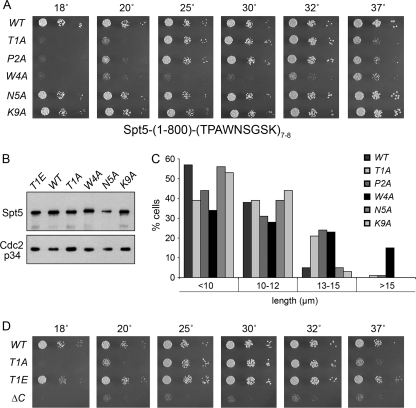
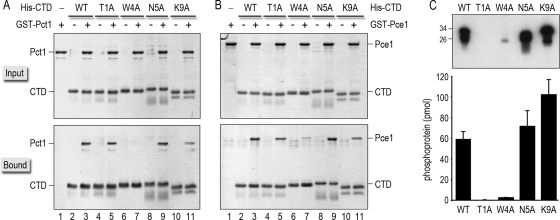
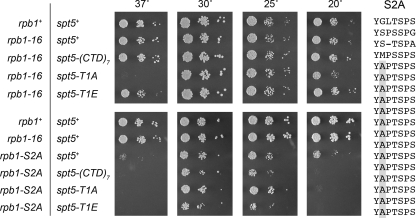
An interaction network connecting mRNA capping enzymes, the RNA polymerase II (Pol II) carboxyl-terminal domain (CTD), elongation factor Spt5, and the Cdk7 and Cdk9 protein kinases is thought to comprise a transcription elongation checkpoint. A crux of this network is Spt5, which regulates early transcription elongation and has an imputed role in pre-mRNA processing via its physical association with capping enzymes. Schizosaccharomyces pombe Spt5 has a distinctive CTD composed of tandem nonapeptide repeats of the consensus sequence (1)TPAWNSGSK(9). The Spt5 CTD binds the capping enzymes and is a substrate for threonine phosphorylation by the Cdk9 kinase. Here we report that deletion of the S. pombe Spt5 CTD results in slow growth and aberrant cell morphology. The severity of the spt5-DeltaCTD phenotype is exacerbated by truncation of the Pol II CTD and ameliorated by overexpression of the capping enzymes RNA triphosphatase and RNA guanylyltransferase. These results suggest that the Spt5 and Pol II CTDs play functionally overlapping roles in capping enzyme recruitment. We probed structure-activity relations of the Spt5 CTD by alanine scanning of the consensus nonapeptide. The T1A change abolished CTD phosphorylation by Cdk9 but did not affect CTD binding to the capping enzymes. The T1A and P2A mutations elicited cold-sensitive (cs) and temperature-sensitive (ts) growth defects and conferred sensitivity to growth inhibition by 6-azauracil that was exacerbated by partial truncations of the Pol II CTD. The T1A phenotypes were rescued by a phosphomimetic T1E change but not by capping enzyme overexpression. These results imply a positive role for Spt5 CTD phosphorylation in Pol Il transcription elongation in fission yeast, distinct from its capping enzyme interactions. Viability of yeast cells bearing both Spt5 CTD T1A and Pol II CTD S2A mutations heralds that the Cdk9 kinase has an essential target other than Spt5 and Pol II CTD-Ser2.
Figures


Similar articles
-
How an mRNA capping enzyme reads distinct RNA polymerase II and Spt5 CTD phosphorylation codes.Genes Dev. 2014 Jun 15;28(12):1323-36. doi: 10.1101/gad.242768.114. Genes Dev. 2014. PMID: 24939935 Free PMC article.
-
Interactions between fission yeast mRNA capping enzymes and elongation factor Spt5.J Biol Chem. 2002 May 31;277(22):19639-48. doi: 10.1074/jbc.M200015200. Epub 2002 Mar 13. J Biol Chem. 2002. PMID: 11893740
-
Separate domains of fission yeast Cdk9 (P-TEFb) are required for capping enzyme recruitment and primed (Ser7-phosphorylated) Rpb1 carboxyl-terminal domain substrate recognition.Mol Cell Biol. 2012 Jul;32(13):2372-83. doi: 10.1128/MCB.06657-11. Epub 2012 Apr 16. Mol Cell Biol. 2012. PMID: 22508988 Free PMC article.
-
The CDK9-SPT5 Axis in Control of Transcription Elongation by RNAPII.J Mol Biol. 2025 Jan 1;437(1):168746. doi: 10.1016/j.jmb.2024.168746. Epub 2024 Aug 13. J Mol Biol. 2025. PMID: 39147127 Review.
-
The pleiotropic roles of SPT5 in transcription.Transcription. 2022 Feb-Jun;13(1-3):53-69. doi: 10.1080/21541264.2022.2103366. Epub 2022 Jul 25. Transcription. 2022. PMID: 35876486 Free PMC article. Review.
Cited by
-
A positive feedback loop links opposing functions of P-TEFb/Cdk9 and histone H2B ubiquitylation to regulate transcript elongation in fission yeast.PLoS Genet. 2012;8(8):e1002822. doi: 10.1371/journal.pgen.1002822. Epub 2012 Aug 2. PLoS Genet. 2012. PMID: 22876190 Free PMC article.
-
Fission yeast RNA triphosphatase reads an Spt5 CTD code.RNA. 2015 Jan;21(1):113-23. doi: 10.1261/rna.048181.114. Epub 2014 Nov 20. RNA. 2015. PMID: 25414009 Free PMC article.
-
Redundant pathways for removal of defective RNA polymerase II complexes at a promoter-proximal pause checkpoint.Mol Cell. 2024 Dec 19;84(24):4790-4807.e11. doi: 10.1016/j.molcel.2024.10.012. Epub 2024 Nov 5. Mol Cell. 2024. PMID: 39504960
-
Punctuation and syntax of the RNA polymerase II CTD code in fission yeast.Proc Natl Acad Sci U S A. 2012 Oct 30;109(44):18024-9. doi: 10.1073/pnas.1208995109. Epub 2012 Oct 15. Proc Natl Acad Sci U S A. 2012. PMID: 23071310 Free PMC article.
-
The PAF complex and Prf1/Rtf1 delineate distinct Cdk9-dependent pathways regulating transcription elongation in fission yeast.PLoS Genet. 2013;9(12):e1004029. doi: 10.1371/journal.pgen.1004029. Epub 2013 Dec 26. PLoS Genet. 2013. PMID: 24385927 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous