

Vasoactive intestinal peptide induces cell cycle arrest and regulatory functions in human T cells at multiple levels
- PMID: 20231362
- PMCID: PMC2863702
- DOI: 10.1128/MCB.01282-09
Vasoactive intestinal peptide induces cell cycle arrest and regulatory functions in human T cells at multiple levels
Abstract
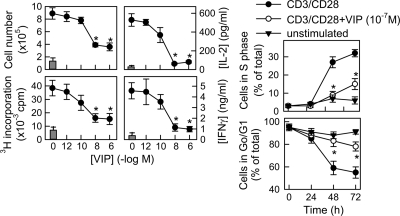
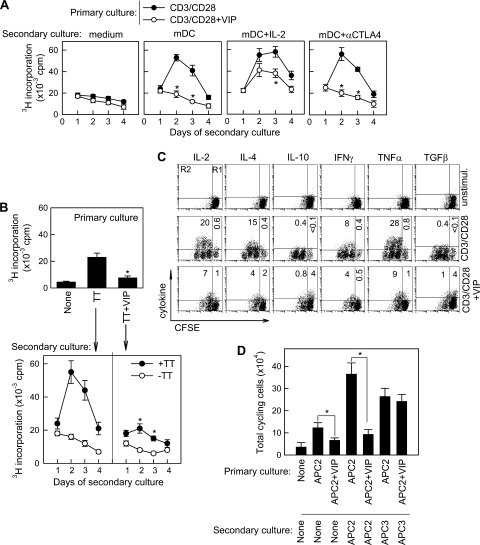
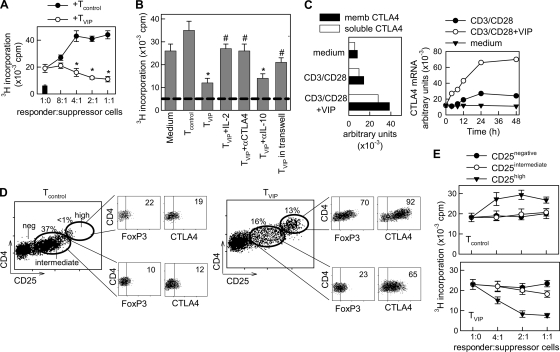
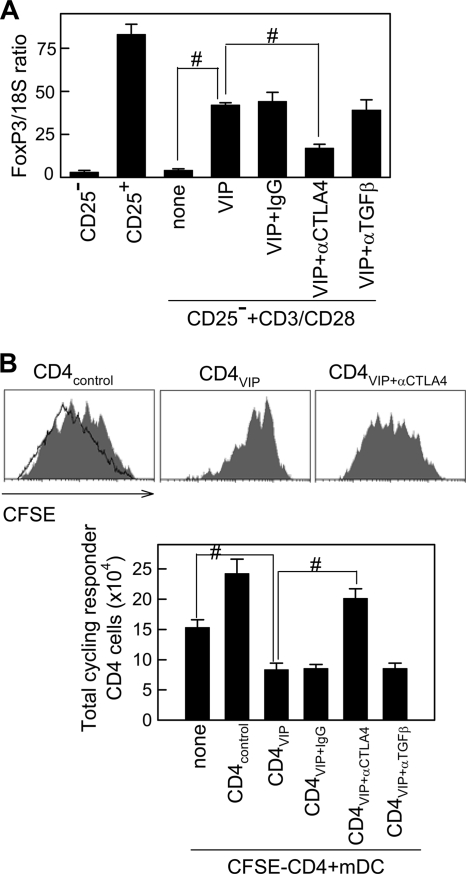
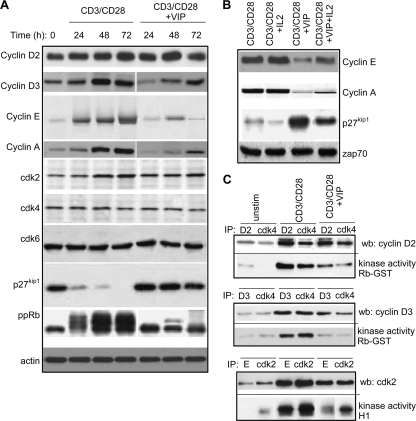
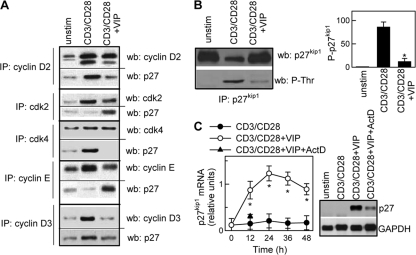
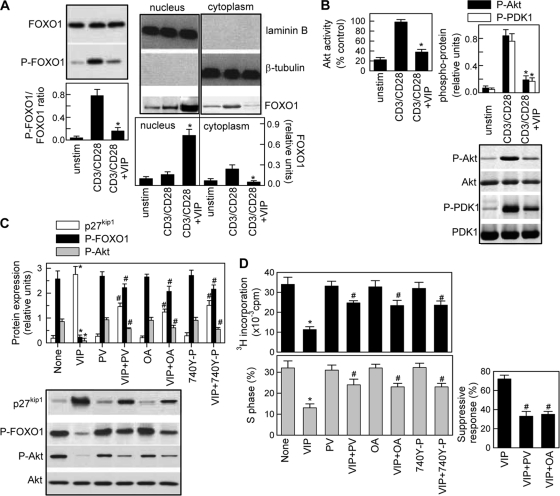
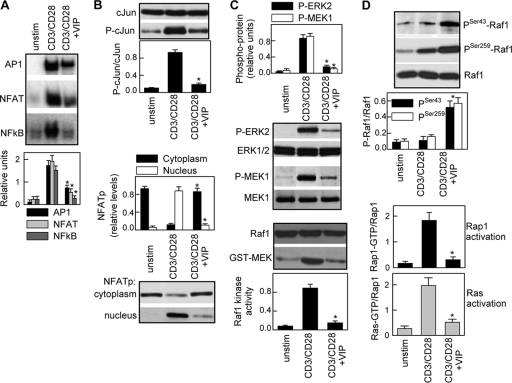
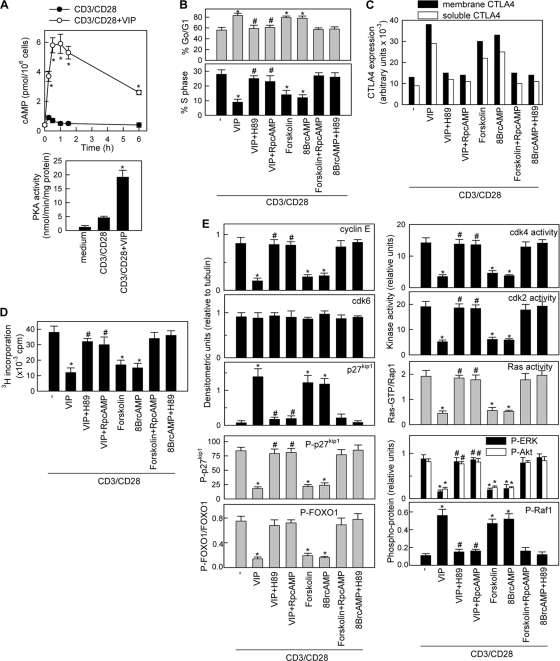
Vasoactive intestinal peptide (VIP) is a potent anti-inflammatory neuropeptide that, by inhibiting Th1-driven responses and inducing the emergence of regulatory T cells (T(reg)), has been proven successful in the induction of tolerance in various experimental models of autoimmune disorders. Here, we investigate the molecular mechanisms involved in VIP-induced tolerance. VIP treatment in the presence of T-cell receptor (TCR) signaling and CD28 costimulation induced cell cycle arrest in human T cells. VIP blocked G(1)/S transition and inhibited the synthesis of cyclins D3 and E and the activation of the cyclin-dependent kinases (CDKs) cdk2 and cdk4. This effect was accompanied by maintenance of threshold levels of the CDK inhibitor p27(kip1) and impairment of phosphatidylinositol 3-kinase (PI3K)-Akt signaling. Inhibition of interleukin 2 (IL-2) transcription and downregulation of signaling through NFAT, AP-1, and Ras-Raf paralleled the VIP-induced cell cycle arrest. Noteworthy from a functional point of view is the fact that VIP-treated T cells show a regulatory phenotype characterized by high expression of CD25, cytotoxic-T-lymphocyte-associated protein 4 (CTLA4), and Forkhead box protein 3 (FoxP3) and potent suppressive activities against effector T cells. CTLA4 appears to be critically involved in the generation and suppressive activities of VIP-induced T(reg). Finally, cyclic AMP (cAMP) and protein kinase A (PKA) activation seems to mediate the VIP-induced cell cycle arrest and T(reg) generation.
Figures
Similar articles
-
CTLA-4-Mediated inhibition of early events of T cell proliferation.J Immunol. 1999 May 15;162(10):5813-20. J Immunol. 1999. PMID: 10229815
-
Induction of alloantigen-specific human T regulatory cells by vasoactive intestinal peptide.J Immunol. 2009 Oct 1;183(7):4346-59. doi: 10.4049/jimmunol.0900400. Epub 2009 Sep 4. J Immunol. 2009. PMID: 19734220
-
Role of bone marrow stromal cells in the generation of human CD8+ regulatory T cells.Hum Immunol. 2008 Nov;69(11):755-9. doi: 10.1016/j.humimm.2008.08.278. Epub 2008 Sep 24. Hum Immunol. 2008. PMID: 18817823 Review.
-
Vav1 promotes T cell cycle progression by linking TCR/CD28 costimulation to FOXO1 and p27kip1 expression.J Immunol. 2006 Oct 15;177(8):5024-31. doi: 10.4049/jimmunol.177.8.5024. J Immunol. 2006. PMID: 17015685
-
Helper T cell anergy: from biochemistry to cancer pathophysiology and therapeutics.J Mol Med (Berl). 2001;78(12):673-83. doi: 10.1007/s001090000180. J Mol Med (Berl). 2001. PMID: 11434720 Review.
Cited by
-
Vasoactive intestinal peptide: a neuropeptide with pleiotropic immune functions.Amino Acids. 2013 Jul;45(1):25-39. doi: 10.1007/s00726-011-1184-8. Epub 2011 Dec 3. Amino Acids. 2013. PMID: 22139413 Free PMC article. Review.
-
Extracellular vesicles derived from T regulatory cells suppress T cell proliferation and prolong allograft survival.Sci Rep. 2017 Sep 14;7(1):11518. doi: 10.1038/s41598-017-08617-3. Sci Rep. 2017. PMID: 28912528 Free PMC article.
-
The Neuropeptides Vasoactive Intestinal Peptide and Pituitary Adenylate Cyclase-Activating Polypeptide Control HIV-1 Infection in Macrophages Through Activation of Protein Kinases A and C.Front Immunol. 2018 Jun 12;9:1336. doi: 10.3389/fimmu.2018.01336. eCollection 2018. Front Immunol. 2018. PMID: 29951068 Free PMC article.
-
AKT isoforms modulate Th1-like Treg generation and function in human autoimmune disease.EMBO Rep. 2016 Aug;17(8):1169-83. doi: 10.15252/embr.201541905. Epub 2016 Jun 16. EMBO Rep. 2016. PMID: 27312110 Free PMC article.
-
Administration of a vasoactive intestinal peptide antagonist enhances the autologous anti-leukemia T cell response in murine models of acute leukemia.Oncoimmunology. 2017 Mar 16;6(5):e1304336. doi: 10.1080/2162402X.2017.1304336. eCollection 2017. Oncoimmunology. 2017. PMID: 28638725 Free PMC article.
References
-
- Alleaume, C., A. Eychène, E. Caigneaux, J. M. Muller, and M. Philippe. 2003. Vasoactive intestinal peptide stimulates proliferation in HT29 human colonic adenocarcinoma cells: concomitant activation of Ras/Rap1-B-Raf-ERK signalling pathway. Neuropeptides 37:98-104. - PubMed
-
- Appleman, L. J., A. A. van Puijenbroek, K. M. Shu, L. M. Nadler, and V. A. Boussiotis. 2002. CD28 costimulation mediates down-regulation of p27kip1 and cell cycle progression by activation of the PI3K/PKB signalling pathway in primary human T cells. J. Immunol. 168:2729-2736. - PubMed
-
- Appleman, L. J., A. Berezovskaya, I. Grass, and V. A. Boussiotis. 2000. CD28 costimulation mediates T cell expansion via IL-2-independent and IL-2-dependent regulation of cell cycle progression. J. Immunol. 164:144-151. - PubMed
-
- Baratelli, F., Y. Lin, L. Zhu, S. C. Yang, N. Heuze-Vourc'h, G. Zeng, K. Reckamp, M. Dohadwala, S. Sharma, and S. M. Dubinett. 2005. Prostaglandin E2 induces FOXP3 gene expression and T regulatory cell function in human CD4+ T cells. J. Immunol. 175:1483-1490. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous