

The gene for aromatase, a rate-limiting enzyme for local estrogen biosynthesis, is a downstream target gene of Runx2 in skeletal tissues
- PMID: 20231365
- PMCID: PMC2863706
- DOI: 10.1128/MCB.00672-09
The gene for aromatase, a rate-limiting enzyme for local estrogen biosynthesis, is a downstream target gene of Runx2 in skeletal tissues
Abstract
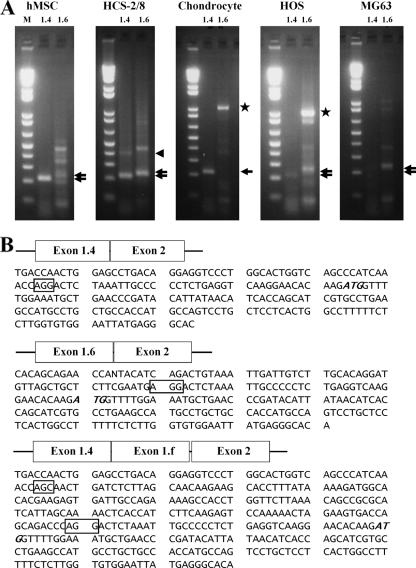
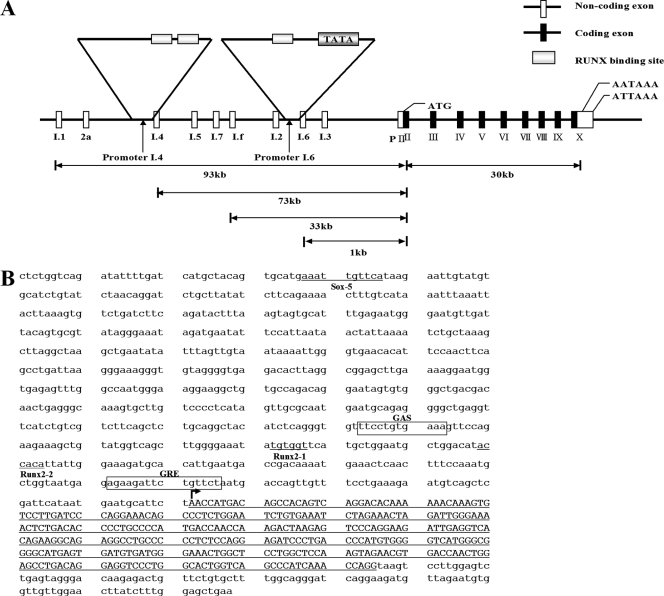
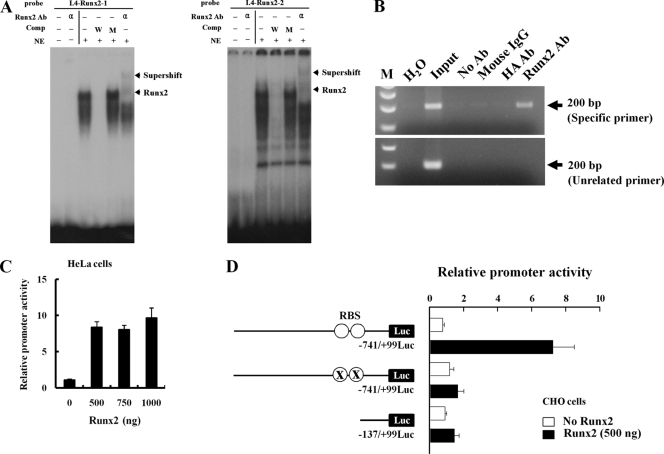
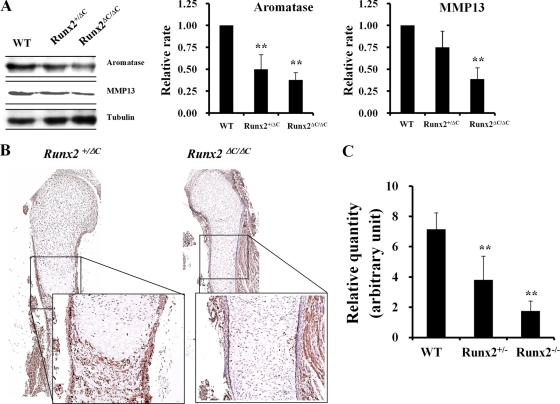
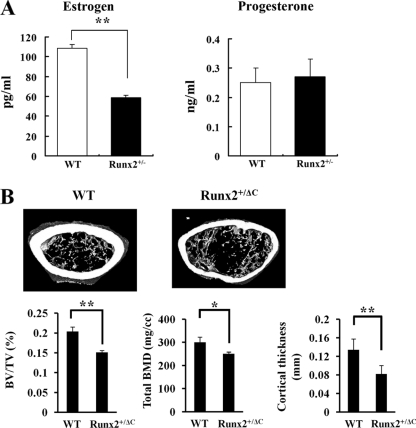
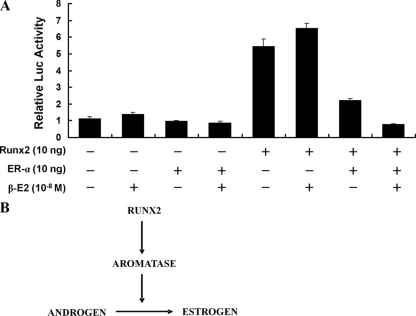
The essential osteoblast-related transcription factor Runx2 and the female steroid hormone estrogen are known to play pivotal roles in bone homeostasis; however, the functional interaction between Runx2- and estrogen-mediated signaling in skeletal tissues is minimally understood. Here we provide evidence that aromatase (CYP19), a rate-limiting enzyme responsible for estrogen biosynthesis in mammals, is transcriptionally regulated by Runx2. Consistent with the presence of multiple Runx2 binding sites, the binding of Runx2 to the aromatase promoter was demonstrated in vitro and confirmed in vivo by chromatin immunoprecipitation assays. The bone-specific aromatase promoter is activated by Runx2, and endogenous aromatase gene expression is upregulated by Runx2 overexpression, establishing the aromatase gene as a target of Runx2. The biological significance of the Runx2 transcriptional control of the aromatase gene is reflected by the enhanced estrogen biosynthesis in response to Runx2 in cultured cells. Reduced in vivo expression of skeletal aromatase gene and low bone mineral density are evident in Runx2 mutant mice. Collectively, these findings uncover a novel link between Runx2-mediated osteoblastogenic processes and the osteoblast-mediated biosynthesis of estrogen as an osteoprotective steroid hormone.
Figures

References
-
- Banerjee, C., L. R. McCabe, J. Y. Choi, S. W. Hiebert, J. L. Stein, G. S. Stein, and J. B. Lian. 1997. Runt homology domain proteins in osteoblast differentiation: AML3/CBFA1 is a major component of a bone-specific complex. J. Cell. Biochem. 66:1-8. - PubMed
-
- Bulun, S. E., S. Sebastian, K. Takayama, T. Suzuki, H. Sasano, and M. Shozu. 2003. The human CYP19 (aromatase P450) gene: update on physiologic roles and genomic organization of promoters. J. Steroid Biochem. Mol. Biol. 86:219-224. - PubMed
-
- Choi, J. Y., J. Pratap, A. Javed, S. K. Zaidi, L. Xing, E. Balint, S. Dalamangas, B. Boyce, A. J. van Wijnen, J. B. Lian, J. L. Stein, S. N. Jones, and G. S. Stein. 2001. Subnuclear targeting of Runx/Cbfa/AML factors is essential for tissue-specific differentiation during embryonic development. Proc. Natl. Acad. Sci. U. S. A. 98:8650-8655. - PMC - PubMed
-
- Conley, A., and M. Hinshelwood. 2001. Mammalian aromatases. Reproduction 121:685-695. - PubMed
-
- Ducy, P., R. Zhang, V. Geoffroy, A. L. Ridall, and G. Karsenty. 1997. Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell 89:747-754. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources