

Prolonged exposure of the HIV-1 gp41 membrane proximal region with L669S substitution
- PMID: 20231447
- PMCID: PMC2851912
- DOI: 10.1073/pnas.0912381107
Prolonged exposure of the HIV-1 gp41 membrane proximal region with L669S substitution
Abstract
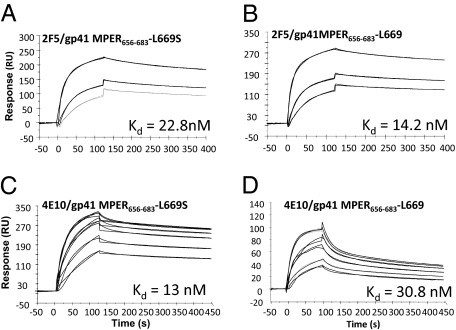
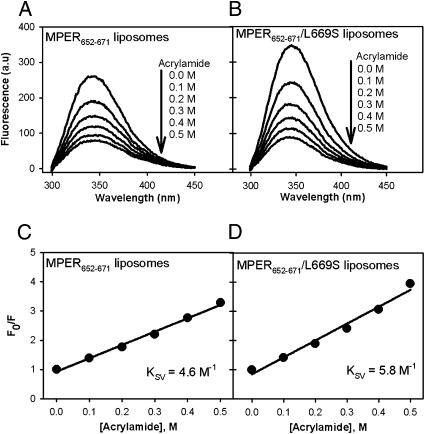
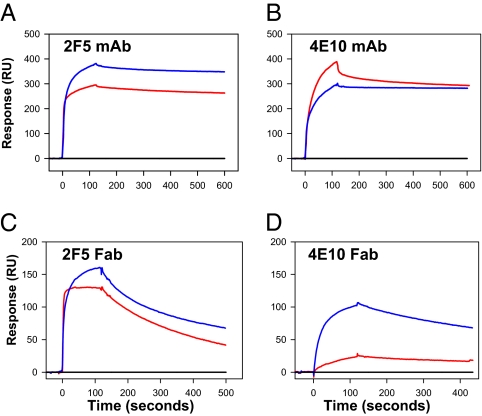
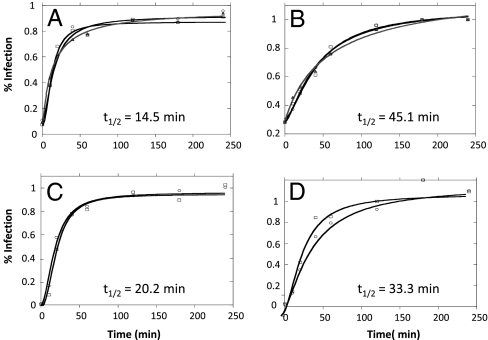
The conserved membrane-proximal external region (MPER) of HIV-1 envelope is a target for the rare broadly neutralizing 2F5, Z13, and 4E10 monoclonal antibodies (mAbs). One strategy to elicit such antibodies is to design an immunogen with increased exposure of the 2F5 and 4E10 mAb epitopes. In this study we characterize a single leucine to serine substitution at position 669 (L669S) in the gp41 Env MPER that confers >250-fold more neutralization sensitivity to 2F5 and 4E10 mAbs than does the wild-type gp41 sequence. On synthetic liposomes, increased solvent exposure of MPER tryptophan residues and stable docking of 2F5 and 4E10 mAbs to mutant MPER peptide liposomes indicate more favorable membrane orientation of MPER neutralizing epitopes with L669S substitution. The time during which virus is sensitive to 2F5 mAb-mediated neutralization is approximately 3-fold longer when the mutation is present. These data suggest that a major contribution to the L669S mutant virus phenotype of enhanced susceptibility to MPER mAbs is prolonged exposure of the MPER neutralizing epitope during viral entry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Anti-human immunodeficiency virus type 1 (HIV-1) antibodies 2F5 and 4E10 require surprisingly few crucial residues in the membrane-proximal external region of glycoprotein gp41 to neutralize HIV-1.J Virol. 2005 Jan;79(2):1252-61. doi: 10.1128/JVI.79.2.1252-1261.2005. J Virol. 2005. PMID: 15613352 Free PMC article.
-
Stable docking of neutralizing human immunodeficiency virus type 1 gp41 membrane-proximal external region monoclonal antibodies 2F5 and 4E10 is dependent on the membrane immersion depth of their epitope regions.J Virol. 2009 Oct;83(19):10211-23. doi: 10.1128/JVI.00571-09. Epub 2009 Jul 29. J Virol. 2009. PMID: 19640992 Free PMC article.
-
An affinity-enhanced neutralizing antibody against the membrane-proximal external region of human immunodeficiency virus type 1 gp41 recognizes an epitope between those of 2F5 and 4E10.J Virol. 2007 Apr;81(8):4033-43. doi: 10.1128/JVI.02588-06. Epub 2007 Feb 7. J Virol. 2007. PMID: 17287272 Free PMC article.
-
Antigp41 membrane proximal external region antibodies and the art of using the membrane for neutralization.Curr Opin HIV AIDS. 2017 May;12(3):250-256. doi: 10.1097/COH.0000000000000364. Curr Opin HIV AIDS. 2017. PMID: 28422789 Review.
-
A new paradigm in molecular recognition? Specific antibody binding to membrane-inserted HIV-1 epitopes.J Mol Recognit. 2011 Jul-Aug;24(4):642-6. doi: 10.1002/jmr.1092. J Mol Recognit. 2011. PMID: 21584875 Review.
Cited by
-
Isotype modulates epitope specificity, affinity, and antiviral activities of anti-HIV-1 human broadly neutralizing 2F5 antibody.Proc Natl Acad Sci U S A. 2012 Jul 31;109(31):12680-5. doi: 10.1073/pnas.1200024109. Epub 2012 Jun 20. Proc Natl Acad Sci U S A. 2012. PMID: 22723360 Free PMC article.
-
Structure of HIV-1 gp41 with its membrane anchors targeted by neutralizing antibodies.Elife. 2021 Apr 19;10:e65005. doi: 10.7554/eLife.65005. Elife. 2021. PMID: 33871352 Free PMC article.
-
Global Increases in Human Immunodeficiency Virus Neutralization Sensitivity Due to Alterations in the Membrane-Proximal External Region of the Envelope Glycoprotein Can Be Minimized by Distant State 1-Stabilizing Changes.J Virol. 2022 Apr 13;96(7):e0187821. doi: 10.1128/jvi.01878-21. Epub 2022 Mar 15. J Virol. 2022. PMID: 35289647 Free PMC article.
-
Modulation of nonneutralizing HIV-1 gp41 responses by an MHC-restricted TH epitope overlapping those of membrane proximal external region broadly neutralizing antibodies.J Immunol. 2014 Feb 15;192(4):1693-706. doi: 10.4049/jimmunol.1302511. Epub 2014 Jan 24. J Immunol. 2014. PMID: 24465011 Free PMC article.
-
Structural Basis for Broad HIV-1 Neutralization by the MPER-Specific Human Broadly Neutralizing Antibody LN01.Cell Host Microbe. 2019 Nov 13;26(5):623-637.e8. doi: 10.1016/j.chom.2019.09.016. Epub 2019 Oct 22. Cell Host Microbe. 2019. PMID: 31653484 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
