

Review
doi: 10.1007/s00018-010-0321-y.
Epub 2010 Mar 16.
Finding the middle ground: how kinetochores power chromosome congression
Affiliations
- PMID: 20232224
- PMCID: PMC2883098
- DOI: 10.1007/s00018-010-0321-y
Item in Clipboard
Review
Finding the middle ground: how kinetochores power chromosome congression
Cell Mol Life Sci.
2010 Jul.
Abstract
Genomic stability requires error-free chromosome segregation during mitosis. Chromosome congression to the spindle equator precedes chromosome segregation in anaphase and is a hallmark of metazoan mitosis. Here we review the current knowledge and concepts on the processes that underlie chromosome congression, including initial attachment to spindle microtubules, biorientation, and movements, from the perspective of the kinetochore.
Figures
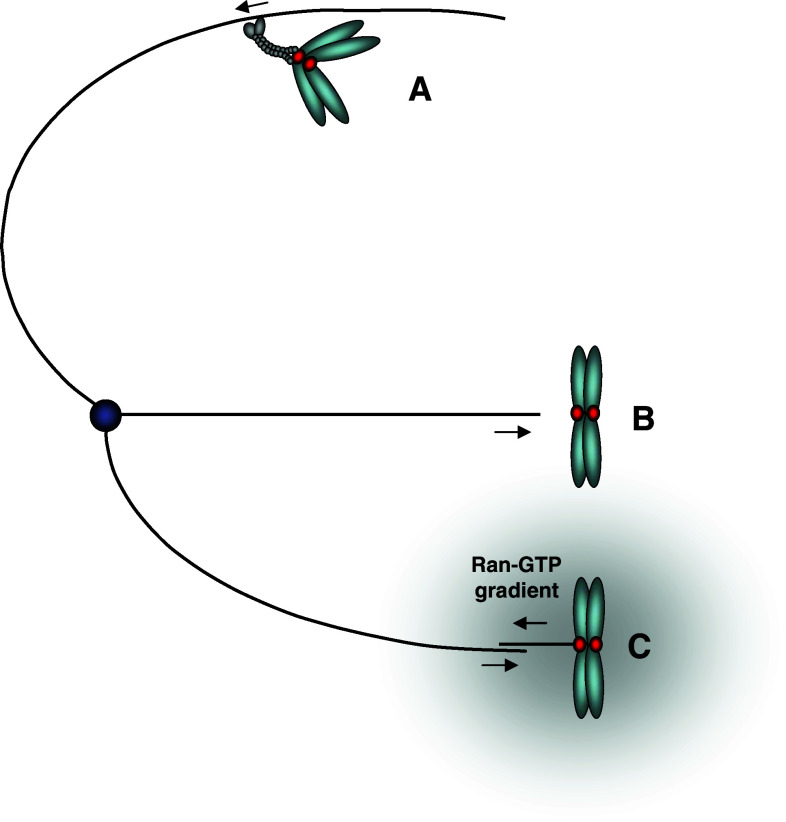
Schematic depicting how kinetochores can initially interact with microtubules. a Lateral attachment to an astral microtubule and subsequent poleward transport by dynein motors. b Direct end-on attachment to the kinetochore. c Interaction between kinetochore-nucleated microtubules (aided by the chromosomal Ran-GTP gradient) and microtubules emanating from the pole. Note that this might be an oversimplification, since the precise relationship between kinetochore-mediated microtubule nucleation and the more general Ran-GTP chromatin-induced microtubule nucleation is until now unclear
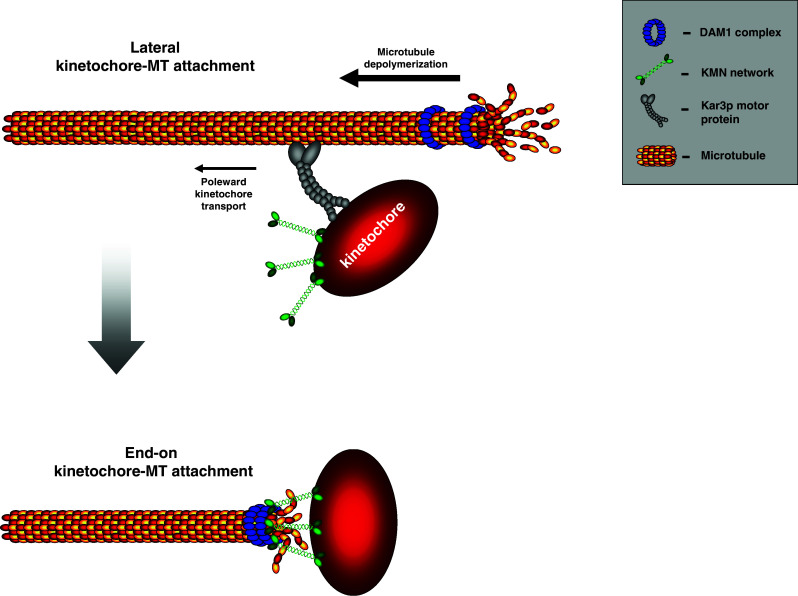
The postulated mechanism for the conversion of lateral attachments into end-on attachments in Saccharomyces cerevisiae. Mono-oriented chromosomes are transported poleward by the Kar3p motor protein (dynein in metazoans). A depolymerizing microtubule accumulates the Dam1 complex (which is proposed to exist as a ring) around the plus-end. When the fast depolymerizing microtubule passes the kinetochore, the Dam1 and the Ndc80 complex within the KMN network will interact to generate an end-on attachment. In metazoans, no clear Dam1 ortholog or plus-end tracking ring structure has been identified. The Ndc80 complex has been shown to track microtubule plus-ends and therefore this interaction may occur directly or through an unknown intermediate binding partner
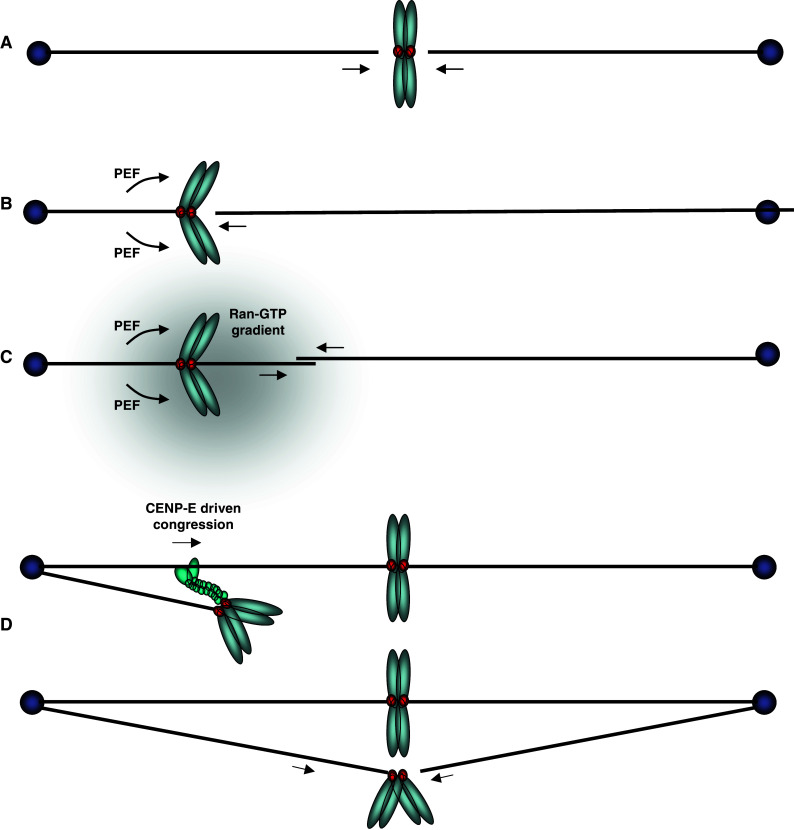
The different routes to kinetochore biorientation. a Direct simultaneous end-on attachment of sister kinetochores with microtubules from both poles. b The mono-oriented sister chromosomes are pushed in an anti-poleward manner by PEF to increase the probability that the opposing sister kinetochore can make a direct end-on attachment with a microtubule from the opposite pole. c The chromosomal Ran-GTP gradient assists microtubule nucleation from the kinetochore, which may then bind directly to microtubules from the distal pole. d The mono-oriented chromosome is transported towards the distal pole along an existing kinetochore fiber by the plus-end directed motor, CENP-E, which binds to the unattached sister kinetochore. This increases the chance of encountering a microtubule emanating from the distal pole
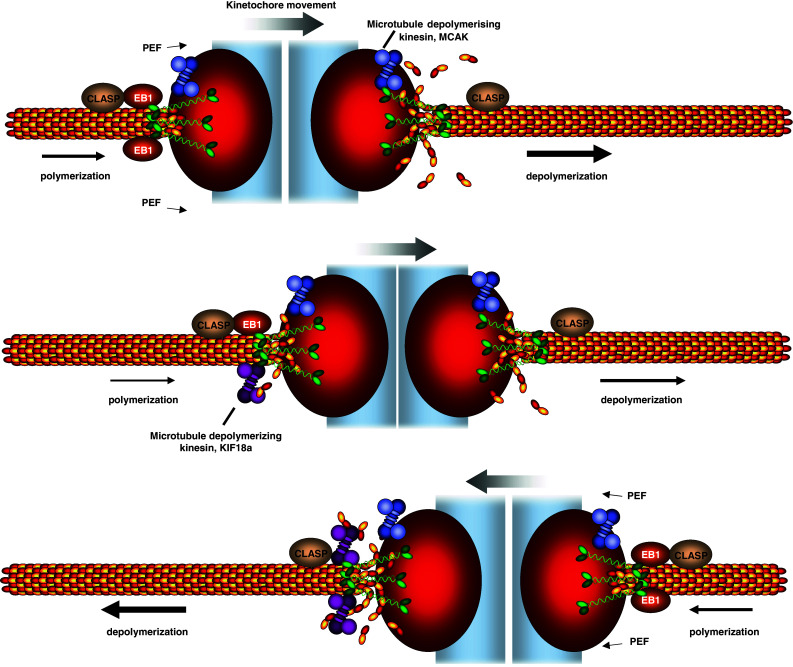
Controlling chromosome movement on the metaphase plate. Bioriented sister kinetochores undergo regular oscillations on the metaphase plate. The direction of chromosome movement is primarily dictated by the pulling forces exerted by plus-end microtubule depolymerization. One proposed regulator of microtubule depolymerization is the microtubule depolymerase MCAK, a kinesin-13. Kinetochore movement is also assisted by microtubule polymerization at the lagging sister kinetochore, which could be promoted by EB1 and requires CLASP. One proposed model for the control of the chromosome oscillations is that the microtubule depolymerase Kif18a accumulates in a length-dependent manner on microtubules, leading to a gradual increase of Kif18a on the lagging kinetochores. As a threshold of Kif18a is achieved, microtubule depolymerization is initiated at the lagging kinetochore, leading to a switch in direction. Note that this schematic is simplified for the purpose of clarity. It depicts one microtubule interacting with the kinetochore, even though in metazoans K-fibers are assembled from up to 25–30 bundled microtubules, raising the question as to how these multiple microtubules are coordinated with each other. Moreover, CLASP and MCAK are known to have more than one attachment site (Clip-170 on microtubule plus-ends and CENP-E on kinetochores for CLASP; inner centromere and inner kinetochore for MCAK [99, 175, 176]) implying a further level of regulation
Similar articles
-
Kinetochore dynein generates a poleward pulling force to facilitate congression and full chromosome alignment.Cell Res. 2007 Aug;17(8):701-12. doi: 10.1038/cr.2007.65. Cell Res. 2007. PMID: 17680027
-
Leaving no-one behind: how CENP-E facilitates chromosome alignment.Essays Biochem. 2020 Sep 4;64(2):313-324. doi: 10.1042/EBC20190073. Essays Biochem. 2020. PMID: 32347304 Free PMC article. Review.
-
Mechanisms of chromosome biorientation and bipolar spindle assembly analyzed by computational modeling.Elife. 2020 Feb 13;9:e48787. doi: 10.7554/eLife.48787. Elife. 2020. PMID: 32053104 Free PMC article.
-
Merotelic kinetochores in mammalian tissue cells.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):553-68. doi: 10.1098/rstb.2004.1610. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897180 Free PMC article. Review.
-
Chromosome congression in the absence of kinetochore fibres.Nat Cell Biol. 2009 Jul;11(7):832-8. doi: 10.1038/ncb1890. Epub 2009 Jun 14. Nat Cell Biol. 2009. PMID: 19525938 Free PMC article.
Cited by
-
Evolutionarily conserved protein ERH controls CENP-E mRNA splicing and is required for the survival of KRAS mutant cancer cells.Proc Natl Acad Sci U S A. 2012 Dec 26;109(52):E3659-67. doi: 10.1073/pnas.1207673110. Epub 2012 Dec 10. Proc Natl Acad Sci U S A. 2012. PMID: 23236152 Free PMC article.
-
Springs, clutches and motors: driving forward kinetochore mechanism by modelling.Chromosome Res. 2011 Apr;19(3):409-21. doi: 10.1007/s10577-011-9191-x. Chromosome Res. 2011. PMID: 21331796 Free PMC article. Review.
-
ERH overexpression is associated with decreased cell migration and invasion and a good prognosis in gastric cancer.Transl Cancer Res. 2020 Sep;9(9):5281-5291. doi: 10.21037/tcr-20-1498. Transl Cancer Res. 2020. PMID: 35117894 Free PMC article.
-
Mitotic chromosome alignment ensures mitotic fidelity by promoting interchromosomal compaction during anaphase.J Cell Biol. 2019 Apr 1;218(4):1148-1163. doi: 10.1083/jcb.201807228. Epub 2019 Feb 7. J Cell Biol. 2019. PMID: 30733233 Free PMC article.
-
The enigmatic ERH protein: its role in cell cycle, RNA splicing and cancer.Protein Cell. 2013 Nov;4(11):807-12. doi: 10.1007/s13238-013-3056-3. Protein Cell. 2013. PMID: 24078386 Free PMC article. Review.
References
-
- Waldeyer HWG. Über Karyokinese und ihre Beziehungen zu den Befruchtungsvorgängen. Archiv für mikroskopische Anatomie und Entwicklungsmechanik. 1888;32:1–122.
-
- Flemming W. Beitrage zur kenntnis der zelle und ihrer lebenserscheinungen. Arch Mikrosk Anat. 1879;18:302–436. doi: 10.1007/BF02956386. - DOI
-
- Flemming W (1882) Zellsubstanz, Kern und Zelltheilung. F.C.W. Vogel. Leipzig
-
- Boveri T. Über mehrpolige Mitosen als Mittel zur Analyse des Zellkerns. Verh. d. phys.-med. Ges. 1902;35:67–90.
-
- Sutton WS. The chromosomes in heredity. Biol Bull. 1903;4:231–251. doi: 10.2307/1535741. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
